Митохондрии содержатся не только в клетках растений, но также и в клетках животных и грибов. Эти органеллы более универсальны, чем пластиды. Впервые ДНК в митохондриях были открыты в 1963 году (М. Наас) сразу же после открытия ДНК в пластидах. Несмотря на сходство функций и структуры митохондрий всех трех царств эукариот, их генетическая организация достаточно сильно различается, поэтому обычно организацию геномов митохондрий у этих царств рассматривают отдельно, выявляя при этом общие черты организации генома.
Физико-химический состав ДНК митохондрий у различных царств различен. У растений он довольно постоянен: от 45 до 47 % ДНК состоит из ГЦ-пар. У животных и грибов -- варьирует более значительно: от 21 до 50 % ГЦ-пар.
У многоклеточных животных размеры генома митохондрий колеблются от 14.5 до 19.5 т.п.н. Практически, это всегда одна кольцевая молекула ДНК. Например, ДНК митохондрий человека -- кольцевая молекула размером 16 569 пар нуклеотидов. Этот размер можно выразить и в других единицах -- в виде молекулярной массы -- 10 6 дальтон или в виде длины контура молекулы -- 5 мкм. Первичная структура этой молекулы полностью определена. В митохондриях содержится собственный аппарат трансляции -- т.е. собственные рибосомы 70S, похожие на хлоропластные или прокариотические и состоящие из двух субъединиц, собственные матричные РНК, необходимые ферменты и белковые факторы. В их геноме закодированы 12S- и 16S - рибосомальные РНК, а так же 22 транспортные РНК. Кроме того, митохондриальная ДНК кодирует 13 полипептидов, из которых 12 идентифицированы. Все кодирующие последовательности расположены прямо друг за другом. В крайнем случае, они разделены лишь несколькими нуклеотидами. Некодирующие последовательности, т.е. интроны отсутствуют. Вслед за кодирующей последовательностью почти всегда находится ген транспортной РНК. Например, порядок таков: транспортная РНК фенилаланина -- ген 12S рибосомальной РНК -- транспортная РНК валина -- ген 16S рибосомальной РНК -- транспортная РНК лейцина и т.д. Такой порядок характерен не только для митохондрий человека, он очень консервативен и характерен для всех животных: дрозофилы, быка, мыши, птиц, рептилий и др. животных.
Большая часть генов расположена в тяжелой цепи, в легкой цепи только гены восьми транспортных РНК и один структурный ген. Таким образом, в отличие от всех других геномов, в геноме митохондрий обе цепи смысловые.
Хотя порядок генов у митохондрий животных и одинаков, выяснено, что сами гены обладают различной консервативностью. Наиболее вариабельна последовательность нуклеотидов участка начала репликации и ряд структурных генов. Наиболее консервативные последовательности расположены в генах рибосомальных РНК и некоторых структурных генах, в том числе в кодирующей последовательности АТФ-азы.
Следует отметить, что универсальность генетического кода нарушена в геноме митохондрий. Например, митохондрии человека используют триплет AUA в качестве кодона для метионина, а не для изолейцина, как у всех, а триплет UGA, используемый в стандартном генетическом словаре как терминирующий кодон, у митохондрий кодирует триптофан.
В целом митохондриальная ДНК человека выглядит так же, как и других млекопитающих: мыши и быка. Несмотря на то, что это далеко не близкие виды -- размеры их митохондриальных ДНК довольно близки между собой: 16 569; 16 295; и 16 338 пар оснований, соответственно. Гены транспортной РНК разделяют некоторые смысловые гены. Наиболее важные из структурных генов -- гены цитохромоксидазы, NADH-дегидрогеназы, цитохром-С оксидоредуктазы и АТФ-синтетазы (рис. 4).
На карте митохондриального генома человека, кроме генов показано и пять хорошо известных болезней человека, наследующихся по материнской линии и вызванных мутациями в митохондриальном геноме.
Так, например, болезнь Лебера -- атрофия зрительного нерва -- вызвана мутацией в гене NADH дегидрогеназы. Эта же болезнь может быть вызвана и мутацией в гене цитохрома b и других локусов. Всего известно нарушение четырех локусов, способных вызвать тот же мутантный фенотип. Кроме того, на этой же карте показано еще четыре болезни, связанные с дефектами мозга, мышц, сердца, почек и печени. Все эти болезни наследуются по материнской линии, и если мать имеет не только дефектные, но и нормальные митохондриальные ДНК и митохондрии, то, происходит сортировка мутантных и нормальных органелл, и потомство может иметь и те, и другие органеллы в различных пропорциях, и мы можем наблюдать также и соматическое расщепление, когда отдельные части тела не будут иметь этих дефектов.
Рис. 4 Структура митохондриального генома млекопитающих, основанная на полном сиквенсе митохондриальной ДНК человека, мыши и быка
Таким образом, небольшой по размерам митохондриальный геном животных может кодировать чрезвычайно важные функции организма и в значительной степени определять его нормальное развитие.
Так же, как и геном пластид, геном митохондрий кодирует только часть митохондриальных полипептидов (табл. 1) и наблюдается феномен двойного кодирования. Например, часть субъединиц АТФ-азного комплекса кодируется ядром, в то время как другая часть -- геномом митохондрий. Большая часть генов, кодирующих рибосомальные миохондриальные РНК и белки, а также ферменты транскрипции и трансляции, кодируется ядром клетки.
Происхождение, то есть были приобретены предками эукариот лишь однажды.
На основании сходства в последовательностях нуклеотидов ДНК ближайшими родственниками митохондрий среди ныне живущих прокариот считают альфа-протеобактерий (в частности, выдвигалась гипотеза, что к митохондриям близки риккетсии). Сравнительный анализ геномов митохондрий показывает, что в ходе эволюции происходило постепенное перемещение генов предков современных митохондрий в ядро клетки. Необъяснимыми с эволюционной точки зрения остаются некоторые особенности митохондриальной ДНК (например, довольно большое число интронов , нетрадиционное использование триплетов и другие). Ввиду ограниченного размера митохондриального генома бо́льшая часть митохондриальных белков кодируется в ядре. При этом бо́льшая часть митохондриальных тРНК кодируются митохондриальным геномом.
Формы и число молекул митохондриальной ДНК
У большинства изученных организмов митохондрии содержат только кольцевые молекулы ДНК, у некоторых растений одновременно присутствуют и кольцевые, и линейные молекулы, а у ряда протистов (например, инфузорий) имеются только линейные молекулы.
У растений каждая митохондрия содержит несколько молекул ДНК разного размера, которые способны к рекомбинации.
При половом размножении митохондрии, как правило, наследуются исключительно по материнской линии, митохондрии сперматозоида обычно разрушаются после оплодотворения. Кроме того, большая часть митохондрий сперматозоида находятся в основании жгутика , которое при оплодотворении иногда теряется. В 1999 году было обнаружено, что митохондрии сперматозоидов помечены убиквитином (белком-меткой, которая приводит к разрушению отцовских митохондрий в зиготе) .
Так как митохондриальная ДНК не является высококонсервативной и имеет высокую скорость мутирования, она является хорошим объектом для изучения филогении (эволюционного родства) живых организмов. Для этого определяют последовательности митохондриальной ДНК у разных видов и сравнивают их при помощи специальных компьютерных программ и получают эволюционное древо для изученных видов. Исследование митохондриальных ДНК собак позволило проследить происхождение собак от диких волков . Исследование митохондриальной ДНК в популяциях человека позволило вычислить «митохондриальную Еву », гипотетическую прародительницу всех живущих в настоящее время людей.
Наследование по отцовской линии
Для некоторых видов показана передача митохондриальной ДНК по мужской линии, например, у мидий . Наследование митохондрий по отцовской линии также описано для некоторых насекомых, например, для дрозофилы , медоносных пчел и цикад .
Существуют также данные о митохондриальном наследовании по мужской линии у млекопитающих. Описаны случаи такого наследования для мышей, при этом митохондрии, полученные от самца, впоследствии отторгаются. Такое явление показано для овец и клонированного крупного рогатого скота.
Наследование по отцовской линии у людей
До недавнего времени считалось, что митохондрии человека наследуются только по материнской линии. Был известен лишь один-единственный случай пациента, у которого в 2002 году достоверно обнаружили отцовскую митохондриальную ДНК .
Лишь недавнее исследование 2018 года показало, что митохондриальная ДНК человека иногда всё же может передаваться и по отцовской линии. Небольшое количество митохондрий отца может попасть в яйцеклетку матери вместе с цитоплазмой сперматозоида, но, как правило, отцовские митохондрии после этого из зиготы исчезают. Однако, было обнаружено, что у некоторых людей существует «мутация, которая помогает выживать митохондриям отца» .
Геном митохондрий
У млекопитающих каждая молекула мтДНК содержит 15000-17000 пар оснований (у человека 16565 пар нуклеотидов - исследование закончено в 1981 году, по другому источнику 16569 пар ) и содержит 37 генов - 13 кодируют белки, 22 - гены тРНК , 2 - рРНК (по одному гену для 12S и 16S рРНК). Другие многоклеточные животные имеют схожий набор митохондриальных генов, хотя некоторые гены могут иногда отсутствовать. Генный состав мтДНК разных видов растений, грибов и особенно протистов различается более значительно. Так, у жгутиконосца-якобиды Reclinomonas americana найден наиболее полный из известных митохондриальных геномов: он содержит 97 генов , в том числе 62 гена, кодирующих белки (27 рибосомальных белков, 23 белка, участвующих в работе электрон-транспортной цепи и в окислительном фосфорилировании , а также субъединицы РНК-полимеразы).
Один из наиболее маленьких митохондриальных геномов имеет малярийный плазмодий (около 6.000 п.о., содержит два гена рРНК и три гена, кодирующих белки).
Недавно открытые рудиментарные митохондрии (митосомы) некоторых протистов (дизентерийной амёбы , микроспоридий и лямблий) не содержат ДНК.
Митохондриальные геномы различных видов грибов содержат от 19 431 (делящиеся дрожжи Schizosaccharomyces pombe ) до 100 314 (сордариомицет Podospora anserina ) пар нуклеотидов .
Некоторые растения имеют огромные молекулы митохондриальной ДНК (до 25 миллионов пар оснований), при этом содержащие примерно те же гены и в том же количестве, что и меньшие мтДНК. Длина митохондриальной ДНК может широко варьировать даже у растений одного семейства. В митохондриальной ДНК растений имеются некодирующие повторяющиеся последовательности.
Геном человека содержит только по одному промотору на каждую комплементарную цепь ДНК .
Геном митохондрий человека кодирует следующие белки и РНК:
| Белки или РНК | Гены |
| NADH-дегидрогеназа (комплекс I) |
MT-ND1, MT-ND2, MT-ND3, MT-ND4, MT-ND4L, MT-ND5, MT-ND6 |
| Кофермент Q - цитохром c редуктаза/Цитохром b (комплекс III) |
MT-CYB |
| цитохром c оксидаза (комплекс IV) |
MT-CO1, MT-CO2, MT-CO3 |
| АТФ-синтаза | MT-ATP6, MT-ATP8 |
| рРНК | MT-RNR1 (12S), MT-RNR2 (16S) |
| тРНК | MT-TA, MT-TC, MT-TD, MT-TE, MT-TF, MT-TG, MT-TH, MT-TI, MT-TK, MT-TL1 , MT-TL2, MT-TM, MT-TN, MT-TP, MT-TQ, MT-TR, MT-TS1, MT-TS2, MT-TT, MT-TV, MT-TW, MT-TY, MT1X |
Особенности митохондриальной ДНК
Кодирующие последовательности (кодоны) митохондриального генома имеют некоторые отличия от кодирующих последовательностей универсальной ядерной ДНК.
Так, кодон AUA кодирует в митохондриальном геноме метионин (вместо изолейцина в ядерной ДНК), кодоны AGA и AGG - терминаторные кодоны (в ядерной ДНК кодируют аргинин), кодон UGA в митохондриальном геноме кодирует триптофан .
Если говорить точнее, то речь идёт не о митохондриальной ДНК, а о мРНК , которая списывается (транскрибируется) с этой ДНК перед началом синтеза белка. Буква U в обозначении кодона обозначает уридин , который при транскрипции гена в РНК заменяет тимин .
Количество генов тРНК (22 гена) меньше, чем в ядерном геноме с его 32 генами тРНК .
В человеческом митохондриальном геноме информация настолько сконцентрирована, что в последовательностях, кодирующих мРНК, как правило, частично удалены нуклеотиды, соответствующие 3"-концевым терминаторным кодонам .
Применение
Кроме использования при построении различных филогенетических теорий, изучение митохондриального генома - основной инструмент при проведении идентификации . Возможность идентификации связана с существующими в митохондриальном геноме человека групповыми и даже индивидуальными различиями.
Последовательность участка гена субъединицы I цитохром с-оксидазы, кодируемого в митохондриальной ДНК, широко используется в проектах, связанных с ДНК-баркодированием животных - определением принадлежности организма к тому или иному таксону на основе коротких маркеров в его ДНК . Для баркодирования растений используется преимущественно комбинация двух маркёров в пластидной ДНК .
Группа Шухрата Миталипова из центра эмбриональных клеток и генной терапии Орегонского университета разработала метод замены митохондриальной ДНК для лечения наследственных митохондриальных заболеваний. Сейчас в Великобритании начаты клинические испытания этого метода, получившего неофициальное название «3-parent baby technique» - «ребенок от трех родителей». Известно также о рождении в результате этой процедуры ребенка в Мексике .
Примечания
- Джинкс Д., Нехромосомная наследственность, пер. с англ., М., 1966; Сэджер Р., Гены вне хромосом, в кн.: Молекулы и клетки, пер. с англ., М., 1966.
- Nass, M.M. & Nass, S. (1963 at the Wenner-Gren Institute for Experimental Biology, Stockholm University, Stockholm , Sweden): Intramitochondrial Fibers with DNA characteristics (PDF). In: J. Cell. Biol. Bd. 19, S. 593-629. PMID 14086138
- Ellen Haslbrunner, Hans Tuppy and Gottfried Schatz (1964 at the Institut for Biochemistry at the Medical Faculty of the University of Vienna in Vienna , Австрия): «Deoxyribonucleic Acid Associated with Yeast Mitochondria» (PDF) Biochem. Biophys. Res. Commun. 15, 127-132.
- Iborra F. J., Kimura H., Cook P. R. The functional organization of mitochondrial genomes in human cells (англ.) // BMC Biol. (англ.) русск. : journal. - 2004. - Vol. 2 . - P. 9 . - DOI :10.1186/1741-7007-2-9 . - PMID 15157274 .
- Дымшиц Г. М. Сюрпризы митохондриального генома. Природа, 2002, N 6
- Wiesner R. J., Ruegg J. C., Morano I. Counting target molecules by exponential polymerase chain reaction, copy number of mitochondrial DNA in rat tissues (англ.) // Biochim Biophys Acta. (англ.) русск. : journal. - 1992. - Vol. 183 . - P. 553-559 . - PMID 1550563 .
- doi:10.1016/j.exppara.2006.04.005 (недоступная ссылка)
- Alexeyev, Mikhail F.; LeDoux, Susan P.; Wilson, Glenn L. Mitochondrial DNA and aging (неопр.) // Clinical Science. - 2004. - July (т. 107 , № 4 ). - С. 355-364 . - DOI :10.1042/CS20040148 . - PMID 15279618 .
- Ченцов Ю. С. Общая цитология. - 3-е изд. - МГУ, 1995. - 384 с. - ISBN 5-211-03055-9 .
- Sutovsky, P., et. al. Ubiquitin tag for sperm mitochondria (англ.) // Nature . - Nov. 25, 1999. - Vol. 402 . - P. 371-372 . - DOI :10.1038/46466 . - PMID 10586873 . Discussed in
- Vilà C., Savolainen P., Maldonado J. E., and Amorin I. R. Multiple and Ancient Origins of the Domestic Dog (англ.) // Science : journal. - 1997. - 13 June (vol. 276 ). - P. 1687-1689 . - ISSN 0036-8075 . - DOI :10.1126/science.276.5319.1687 . - PMID 9180076 .
- Hoeh W. R., Blakley K. H., Brown W. M. Heteroplasmy suggests limited biparental inheritance of Mytilus mitochondrial DNA (англ.) // Science: journal. - 1991. - Vol. 251 . - P. 1488-1490 . - DOI :10.1126/science.1672472 . - PMID 1672472 .
- Penman, Danny . Mitochondria can be inherited from both parents , NewScientist.com (23 августа 2002). Дата обращения 5 февраля 2008.
- Kondo R., Matsuura E. T., Chigusa S. I. Further observation of paternal transmission of Drosophila mitochondrial DNA by PCR selective amplification method (англ.) // Genet. Res. (англ.) русск. : journal. - 1992. - Vol. 59 , no. 2 . - P. 81-4 . - PMID 1628820 .
- Meusel M. S., Moritz R. F. Transfer of paternal mitochondrial DNA during fertilization of honeybee (Apis mellifera L.) eggs (англ.) // Curr. Genet. : journal. - 1993. - Vol. 24 , no. 6 . - P. 539-543 . - DOI :10.1007/BF00351719 . - PMID 8299176 .
- Fontaine, K. M., Cooley, J. R., Simon, C. Evidence for paternal leakage in hybrid periodical cicadas (Hemiptera: Magicicada spp.) (исп.) // PLoS One. : diario. - 2007. - V. 9 . - P. e892 . - DOI :10.1371/journal.pone.0000892 .
- Gyllensten U., Wharton D., Josefsson A., Wilson A. C. Paternal inheritance of mitochondrial DNA in mice (англ.) // Nature. - 1991. - Vol. 352 , no. 6332 . - P. 255-257 . - DOI :10.1038/352255a0 . - PMID 1857422 .
- Shitara H., Hayashi J. I., Takahama S., Kaneda H., Yonekawa H. Maternal inheritance of mouse mtDNA in interspecific hybrids: segregation of the leaked paternal mtDNA followed by the prevention of subsequent paternal leakage (англ.) // Genetics: journal. - 1998. - Vol. 148 , no. 2 . - P. 851-857 . - PMID 9504930 .
© Г.М.Дымшиц
Сюрпризы митохондриального генома
Г.М. Дымшиц
Григорий Моисеевич Дымшиц, доктор биологических наук, профессор кафедры молекулярной биологии Новосибирского государственного университета, заведующий лабораторией структуры генома Института цитологии и генетики Сибирского отделения РАН. Соавтор и редактор четырех школьных учебников по общей биологии.Со времени обнаружения в митохондриях молекул ДНК прошло четверть века, прежде чем ими заинтересовались не только молекулярные биологи и цитологи, но и генетики, эволюционисты, а также палеонтологи и криминалисты, историки и лингвисты. Такой широкий интерес спровоцировала работа А.Уилсона из Калифорнийского университета. В 1987 г. он опубликовал результаты сравнительного анализа ДНК митохондрий, взятых у 147 представителей разных этносов всех человеческих рас, заселяющих пять континентов. По типу, местоположению и количеству индивидуальных мутаций установили, что все митохондриальные ДНК возникли из одной предковой последовательности нуклеотидов путем дивергенции. В околонаучной прессе вывод этот интерпретировали крайне упрощенно - все человечество произошло от одной женщины, названной митохондриальной Евой (и дочери и сыновья получают митохондрии только от матери), которая жила в Северо-Восточной Африке около 200 тыс. лет назад. Еще через 10 лет удалось расшифровать фрагмент ДНК митохондрий, выделенный из останков неандертальца, и оценить время существования последнего общего предка человека и неандертальца в 500 тыс. лет назад .
Сегодня митохондриальная генетика человека интенсивно развивается как в популяционном, так и в медицинском аспекте. Установлена связь между рядом тяжелых наследственных заболеваний и дефектами в митохондриальных ДНК. Генетические изменения, ассоциированные со старением организма, наиболее выражены в митохондриях. Что же представляет из себя геном митохондрий, отличающийся у человека и других животных от такового у растений, грибов и простейших и по размеру, и по форме, и по генетической емкости? Как работает и как возник митохондриальный геном у разных таксонов? Об этом и пойдет речь в нашей статье.
Митохондрии называют энергетическими станциями клетки. Помимо наружной гладкой мембраны они имеют внутреннюю мембрану, образующую многочисленные складки - кристы. В них встроены белковые компоненты дыхательной цепи - ферменты, участвующие в преобразовании энергии химических связей окисляемых питательных веществ в энергию молекул аденозинтрифосфорной кислоты (АТФ). Такой “конвертируемой валютой” клетка оплачивает все свои энергетические потребности. В клетках зеленых растений помимо митохондрий есть еще и другие энергетические станции - хлоропласты. Они работают на “солнечных батареях”, но тоже образуют АТФ из АДФ и фосфата. Как и митохондрии, хлоропласты - автономно размножающиеся органеллы - также имеют две мембраны и содержат ДНК.
В матриксе митохондрий, кроме ДНК, находятся и собственные рибосомы, по многим характеристикам отличающиеся от эвкариотических рибосом, расположенных на мембранах эндоплазматической сети. Однако на рибосомах митохондрий образуется не более 5% от всех белков, входящих в их состав. БOльшая часть белков, составляющих структурные и функциональные компоненты митохондрий, кодируется ядерным геномом, синтезируется на рибосомах эндоплазматической сети и транспортируется по ее каналам к месту сборки. Таким образом, митохондрии - это результат объединенных усилий двух геномов и двух аппаратов транскрипции и трансляции. Некоторые субъединичные ферменты дыхательной цепи митохондрий состоят из разных полипептидов, часть которых кодируется ядерным, а часть - митохондриальным геномом. Например, ключевой фермент окислительного фосфорилирования - цитохром-с-оксидаза у дрожжей состоит из трех субъединиц, кодируемых и синтезируемых в митохондриях, и четырех, кодируемых в ядре клетки и синтезируемых в цитоплазме. Экспрессией большинства генов митохондрий управляют определенные гены ядер.
Размеры и формы митохондриальных геномов
К настоящему времени прочитано более 100 разных геномов митохондрий. Набор и количество их генов в митохондриальных ДНК, для которых полностью определена последовательность нуклеотидов, сильно различаются у разных видов животных, растений, грибов и простейших. Наибольшее количество генов обнаружено в митохондриальном геноме жгутикового простейшего Rectinomonas americana - 97 генов, включая все кодирующие белок гены, найденные в мтДНК других организмов. У большинства высших животных геном митохондрий содержит 37 генов: 13 для белков дыхательной цепи, 22 для тРНК и два для рРНК (для большой субъединицы рибосом 16S рРНК и для малой 12S рРНК). У растений и простейших, в отличие от животных и большинства грибов, в митохондриальном геноме закодированы и некоторые белки, входящие в состав рибосом этих органелл. Ключевые ферменты матричного полинуклеотидного синтеза, такие как ДНК-полимераза (осуществляющая репликацию митохондриальной ДНК) и РНК-полимераза (транскрибирующая геном митохондрий), зашифрованы в ядре и синтезируются на рибосомах цитоплазмы. Этот факт указывает на относительность автономии митохондрий в сложной иерархии эвкариотической клетки.
Геномы митохондрий разных видов отличаются не только по набору генов, порядку их расположения и экспрессии, но по размеру и форме ДНК. Подавляющее большинство описанных сегодня митохондриальных геномов представляет собой кольцевые суперспирализованные двуцепочечные молекулы ДНК. У некоторых растений наряду с кольцевыми формами имеются и линейные, а у некоторых простейших, например инфузорий, в митохондриях обнаружены только линейные ДНК .
Как правило, в каждой митохондрии содержится несколько копий ее генома. Так, в клетках печени человека около 2 тыс. митохондрий, и в каждой из них - по 10 одинаковых геномов. В фибробластах мыши 500 митохондрий, содержащих по два генома, а в клетках дрожжей S.cerevisiae - до 22 митохондрий, имеющих по четыре генома.
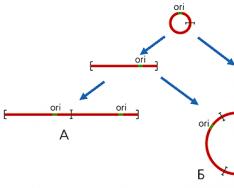
Митохондриальный геном растений, как правило, состоит из нескольких молекул разного размера. Одна из них, “основная хромосома”, содержит большую часть генов, а кольцевые формы меньшей длины, находящиеся в динамическом равновесии как между собой, так и с основной хромосомой, образуются в результате внутри- и межмолекулярной рекомбинации благодаря наличию повторенных последовательностей (рис.1).
Рис 1.
Схема образования кольцевых молекул
ДНК разного размера в митохондриях растений.
Рекомбинация происходит по повторенным участкам (обозначены
синим цветом).

Рис 2.
Схема образования линейных (А), кольцевых
(Б), цепных (В) олигомеров мтДНК.
ori - район начала репликации ДНК.
Размер генома митохондрий разных организмов колеблется от менее 6 тыс. пар нуклеотидов у малярийного плазмодия (в нем, помимо двух генов рРНК, содержится только три гена, кодирующих белки) до сотен тысяч пар нуклеотидов у наземных растений (например, у Arabidopsis thaliana из семейства крестоцветных 366924 пар нуклеотидов). При этом 7-8-кратные различия в размерах мтДНК высших растений обнаруживаются даже в пределах одного семейства. Длина мтДНК позвоночных животных отличается незначительно: у человека - 16569 пар нуклеотидов, у свиньи - 16350, у дельфина - 16330, у шпорцевой лягушки Xenopus laevis - 17533, у карпа - 16400. Эти геномы сходны также и по локализации генов, большинство которых располагаются встык; в ряде случаев они даже перекрываются, обычно на один нуклеотид, так что последний нуклеотид одного гена оказывается первым в следующем. В отличие от позвоночных, у растений, грибов и простейших мтДНК содержат до 80% некодирующих последовательностей. У разных видов порядок генов в геномах митохондрий отличается.
Высокая концентрация активных форм кислорода в митохондриях и слабая система репарации увеличивают частоту мутаций мтДНК по сравнению с ядерной на порядок. Радикалы кислорода служат причиной специфических замен Ц® Т (дезаминирование цитозина) и Г® Т (окислительное повреждение гуанина), вследствие чего, возможно, мтДНК богаты АТ-парами. Кроме того, все мтДНК обладают интересным свойством - они не метилируются, в отличие от ядерных и прокариотических ДНК. Известно, что метилирование (временная химическая модификация нуклеотидной последовательности без нарушения кодирующей функции ДНК) - один из механизмов программируемой инактивации генов .
Репликация и транскрипция ДНК митохондрий млекопитающих
У большинства животных комплементарные цепи в мтДНК значительно различаются по удельной плотности, поскольку содержат неодинаковое количество “тяжелых” пуриновых и “легких” пиримидиновых нуклеотидов. Так они и называются - H (heavy - тяжелая) и L (light - легкая) цепь. В начале репликации молекулы мтДНК образуется так называемая D-петля (от англ. displacement loop - петля смещения). Эта структура, видимая в электронный микроскоп, состоит из двуцепочечного и одноцепочечного (отодвинутой части Н-цепи) участков. Двуцепочечный участок формируется частью L-цепи и комплементарным ей вновь синтезированным фрагментом ДНК длиной 450-650 (в зависимости от вида организма) нуклеотидов, имеющим на 5"-конце рибонуклеотидную затравку, которая соответствует точке начала синтеза Н-цепи (ori H). Синтез L-цепи начинается лишь тогда, когда дочерняя Н-цепь доходит до точки ori L. Это обусловлено тем, что область инициации репликации L-цепи доступна для ферментов синтеза ДНК лишь в одноцепочечном состоянии, а следовательно, только в расплетенной двойной спирали при синтезе Н-цепи. Таким образом, дочерние цепи мтДНК синтезируются непрерывно и асинхронно (рис.3).

Рис 3.
Схема репликации мтДНК млекопитающих.
Сначала формируется D-петля, затем синтезируется дочерняя
Н-цепь,
потом начинается синтез дочерней L-цепи.
В митохондриях общее число молекул с D-петлей значительно превышает число полностью реплицирующихся молекул. Обусловлено это тем, что у D-петли есть дополнительные функции - прикрепление мтДНК к внутренней мембране и инициация транскрипции, поскольку в этом районе локализованы промоторы транскрипции обеих цепей ДНК.
В отличие от большинства эвкариотических генов, которые транскрибируются независимо друг от друга, каждая из цепей мтДНК млекопитающих переписывается с образованием одной молекулы РНК, начинающейся в районе ori H. Помимо этих двух длинных молекул РНК, комплементарных Н- и L-цепям, формируются и более короткие участки Н-цепи, которые начинаются в той же точке и заканчиваются на 3"-конце гена 16S рРНК (рис.4). Таких коротких транскриптов в 10 раз больше, чем длинных. В результате созревания (процессинга) из них образуются 12S рРНК и 16S рРНК, участвующие в формировании митохондриальных рибосом, а также фенилаланиновая и валиновая тРНК. Из длинных транскриптов вырезаются остальные тРНК и образуются транслируемые мРНК, к 3"-концам которых присоединяются полиадениловые последовательности. 5"-концы этих мРНК не кэпируются, что необычно для эвкариот. Сплайсинга (сращивания) не происходит, поскольку ни один из митохондриальных генов млекопитающих не содержит интронов.

Рис 4. Транскрипция мтДНК человека, содержащей 37 генов. Все транскрипты начинают синтезироваться в районе ori H. Рибосомные РНК вырезаются из длинного и короткого транскриптов Н-цепи. тРНК и мРНК образуются в результате процессинга из транскриптов обеих цепей ДНК. Гены тРНК обозначены светло-зеленым цветом.Сюрпризы митохондриального генома
Несмотря на то, что в геномах митохондрий млекопитающих и дрожжей содержится
приблизительно одинаковое количество генов, размеры дрожжевого генома в
4-5 раз больше - около 80 тыс. пар нуклеотидов. Хотя кодирующие последовательности
мтДНК дрожжей высоко гомологичны соответствующим последовательностям у
человека, дрожжевые мРНК дополнительно имеют 5"-лидерную и 3"-некодирующую
области, как и большинство ядерных мРНК. Ряд генов содержит еще и интроны.
Так, в гене box, кодирующем цитохромоксидазу b, имеется два интрона. Из
первичного РНК-транскрипта автокаталитически (без участия каких-либо белков)
вырезается копия большей части первого интрона. Оставшаяся РНК служит матрицей
для образования фермента матуразы, участвующей в сплайсинге. Часть ее аминокислотной
последовательности закодирована в оставшихся копиях интронов. Матураза
вырезает их, разрушая свою собственную мРНК, копии экзонов сшиваются, и
образуется мРНК для цитохромоксидазы b (рис.5). Открытие такого феномена
заставило пересмотреть представление об интронах, как о “ничего не кодирующих
последовательностях”.

Рис 5.
Процессинг (созревание) мРНК цитохромоксидазы
b в митохондриях дрожжей.
На первом этапе сплайсинга образуется мРНК, по которой
синтезируется матураза,
необходимая для второго этапа сплайсинга.
При изучении экспрессии митохондриальных генов Trypanosoma brucei обнаружилось удивительное отклонение от одной из основных аксиом молекулярной биологии, гласящей, что последовательность нуклеотидов в мРНК в точности соответствует таковой в кодирующих участках ДНК. Оказалось, мРНК одной из субъединиц цитохром-с-оксидазы редактируется, т.е. после транскрипции изменяется ее первичная структура - вставляется четыре урацила. В результате образуется новая мРНК, служащая матрицей для синтеза дополнительной субъединицы фермента, последовательность аминокислот в которой не имеет ничего общего с последовательностью, кодируемой нередактированной мРНК (см. таблицу).

Впервые обнаруженное в митохондриях трипаносомы редактирование РНК широко распространено в хлоропластах и митохондриях высших растений. Найдено оно и в соматических клетках млекопитающих, например, в кишечном эпителии человека редактируется мРНК гена аполипопротеина.
Наибольший сюрприз ученым митохондрии преподнесли в 1979 г. До того времени считалось, что генетический код универсален и одни и те же триплеты кодируют одинаковые аминокислоты у бактерий, вирусов, грибов, растений и животных. Английский исследователь Беррел сопоставил структуру одного из митохондриальных генов теленка с последовательностью аминокислот в кодируемой этим геном субъединице цитохромоксидазы. Оказалось, что генетический код митохондрий крупного рогатого скота (как и человека) не просто отличается от универсального, он “идеален”, т.е. подчиняется следующему правилу: “если два кодона имеют два одинаковых нуклеотида, а третьи нуклеотиды принадлежат к одному классу (пуриновых - А, Г, или пиримидиновых - У, Ц), то они кодируют одну и ту же аминокислоту”. В универсальном коде есть два исключения из этого правила: триплет АУА кодирует изолейцин, а кодон АУГ - метионин, в то время как в идеальном коде митохондрий оба эти триплета кодируют метионин; триплет УГГ кодирует лишь триптофан, а триплет УГА - стоп-кодон. В универсальном коде оба отклонения касаются принципиальных моментов синтеза белка: кодон АУГ - инициирующий, а стоп-кодон УГА останавливает синтез полипептида. Идеальный код присущ не всем описанным митохондриям, но ни у одной из них нет универсального кода. Можно сказать, что митохондрии говорят на разных языках, но никогда - на языке ядра.
Как уже говорилось, в митохондриальном геноме позвоночных есть 22 гена тРНК. Каким же образом такой неполный набор обслуживает все 60 кодонов для аминокислот (в идеальном коде из 64 триплетов четыре стоп-кодона, в универсальном - три)? Дело в том, что при синтезе белка в митохондриях упрощены кодон-антикодонные взаимодействия - для узнавания используется два из трех нуклеотидов антикодона. Таким образом, одна тРНК узнает все четыре представителя кодонового семейства, отличающиеся только третьим нуклеотидом. Например, лейциновая тРНК с антикодоном ГАУ встает на рибосоме напротив кодонов ЦУУ, ЦУЦ, ЦУА и ЦУГ, обеспечивая безошибочное включение лейцина в полипептидную цепь. Два других лейциновых кодона УУА и УУГ узнаются тРНК с антикодоном ААУ. В целом, восемь разных молекул тРНК узнают восемь семейств по четыре кодона в каждом, и 14 тРНК узнают разные пары кодонов, каждая из которых шифрует одну аминокислоту.
Важно, что ферменты аминоацил-тРНК-синтетазы, ответственные за присоединение аминокислот к соответствующим тРНК митохондрий, кодируются в ядре клетки и синтезируются на рибосомах эндоплазматической сети. Таким образом, у позвоночных животных все белковые компоненты митохондриального синтеза полипептидов зашифрованы в ядре. При этом синтез белков в митохондриях не подавляется циклогексимидом, блокирующим работу эвкариотических рибосом, но чувствителен к антибиотикам эритромицину и хлорамфениколу, ингибирующим белковый синтез в бактериях. Этот факт служит одним из аргументов в пользу происхождения митохондрий из аэробных бактерий при симбиотическом образовании эвкариотических клеток .
Симбиотическая теория происхождения митохондрий
Гипотезу о происхождении митохондрий и растительных пластид из внутриклеточных бактерий-эндосимбионтов высказал Р.Альтман еще в 1890 г. За век бурного развития биохимии, цитологии, генетики и появившейся полвека назад молекулярной биологии гипотеза переросла в теорию, основанную на большом фактическом материале. Суть ее такова: с появлением фотосинтезирующих бактерий в атмосфере Земли накапливался кислород - побочный продукт их метаболизма. С ростом его концентрации усложнялась жизнь анаэробных гетеротрофов, и часть из них для получения энергии перешла от бескислородного брожения к окислительному фосфорилированию. Такие аэробные гетеротрофы могли с бOльшим КПД, чем анаэробные бактерии, расщеплять органические вещества, образующиеся в результате фотосинтеза. Часть свободно живущих аэробов была захвачена анаэробами, но не “переварена”, а сохранена в качестве энергетических станций, митохондрий. Не стоит рассматривать митохондрии как рабов, взятых в плен, чтобы снабжать молекулами АТФ не способные к дыханию клетки. Они скорее “существа”, еще в протерозое нашедшие для себя и своего потомства лучшее из убежищ, где можно затрачивать наименьшие усилия, не подвергаясь риску быть съеденными.
В пользу симбиотической теории говорят многочисленные факты:
- совпадают размеры и формы митохондрий и свободно живущих аэробных бактерий; те и другие содержат кольцевые молекулы ДНК, не связанные с гистонами (в отличие от линейных ядерных ДНК);Существует представление, что разные царства эвкариот имели разных предков и эндосимбиоз бактерий возникал на разных этапах эволюции живых организмов. Об этом же говорят отличия в строении митохондриальных геномов простейших, грибов, растений и высших животных. Но во всех случаях основная часть генов из промитохондрий попала в ядро, возможно, с помощью мобильных генетических элементов. При включении части генома одного из симбионтов в геном другого интеграция симбионтов становится необратимой.По нуклеотидным последовательностям рибосомные и транспортные РНК митохондрий отличаются от ядерных, демонстрируя при этом удивительное сходство с аналогичными молекулами некоторых аэробных грамотрицательных эубактерий;
Митохондриальные РНК-полимеразы, хотя и кодируются в ядре клетки, ингибируются рифампицином, как и бактериальные, а эвкариотические РНК-полимеразы нечувствительны к этому антибиотику;
Белковый синтез в митохондриях и бактериях подавляется одними и теми же антибиотиками, не влияющими на рибосомы эвкариот;
Липидный состав внутренней мембраны митохондрий и бактериальной плазмалеммы сходен, но сильно отличается от такового наружной мембраны митохондрий, гомологичной другим мембранам эвкариотических клеток;
Кристы, образуемые внутренней митохондриальной мембраной, являются эволюционными аналогами мезосомных мембран многих прокариот;
До сих пор сохранились организмы, имитирующие промежуточные формы на пути к образованию митохондрий из бактерий (примитивная амеба Pelomyxa не имеет митохондрий, но всегда содержит эндосимбиотические бактерии).
Новый геном может создавать метаболические пути, приводящие к образованию полезных продуктов, которые не могут быть синтезированы ни одним из партнеров по отдельности. Так, синтез стероидных гормонов клетками коры надпочечников представляет собой сложную цепь реакций, часть которых происходит в митохондриях, а часть - в эндоплазматической сети . Захватив гены промитохондрий, ядро получило возможность надежно контролировать функции симбионта. В ядре кодируются все белки и синтез липидов наружной мембраны митохондрий, большинство белков матрикса и внутренней мембраны органелл. Самое главное, что ядро кодирует ферменты репликации, транскрипции и трансляции мтДНК, контролируя тем самым рост и размножение митохондрий. Скорость роста партнеров по симбиозу должна быть приблизительно одинаковой. Если хозяин будет расти быстрее, то с каждым его поколением число симбионтов, приходящихся на одну особь, будет уменьшаться, и, в конце концов, появятся потомки, не имеющие митохондрий. Мы знаем, что в каждой клетке организма, размножающегося половым путем, содержится много митохондрий, реплицирующих свои ДНК в промежутке между делениями хозяина. Это служит гарантией того, что каждая из дочерних клеток получит по крайней мере одну копию генома митохондрии.
Цитоплазматическая наследственность
Помимо кодирования ключевых компонентов дыхательной цепи и собственного белоксинтезирующего аппарата, митохондриальный геном в отдельных случаях участвует в формировании некоторых морфологических и физиологических признаков. К таким признакам относятся характерные для ряда видов высших растений синдром NCS (non-chromosomal stripe, нехромосомно кодируемая пятнистость листьев) и цитоплазматическая мужская стерильность (ЦМС), приводящая к нарушению нормального развития пыльцы. Проявление обоих признаков обусловлено изменениями в структуре мтДНК. При ЦМС наблюдаются перестройки геномов митохондрий в результате рекомбинационных событий, ведущих к делециям, дупликациям, инверсиям или инсерциям определенных нуклеотидных последовательностей или целых генов. Такие изменения могут вызывать не только повреждения имеющихся генов, но и появление новых работающих генов.
Цитоплазматическая наследственность, в отличие от ядерной, не подчиняется законам Менделя. Это связано с тем, что у высших животных и растений гаметы от разных полов содержат несопоставимые количества митохондрий. Так, в яйцеклетке мыши имеется 90 тыс. митохондрий, а в сперматозоиде - лишь четыре. Очевидно, что в оплодотворенной яйцеклетке митохондрии преимущественно или только от женской особи, т.е. наследование всех митохондриальных генов материнское. Генетический анализ цитоплазматической наследственности затруднен из-за ядерно-цитоплазматических взаимодействий. В случае цитоплазматической мужской стерильности мутантный митохондриальный геном взаимодействует с определенными генами ядра, рецессивные аллели которых необходимы для развития признака. Доминантные аллели этих генов как в гомо-, так и в гетерозиготном состоянии восстанавливают фертильность растений вне зависимости от состояния митохондриального генома.
Изучение геномов митохондрий, их эволюции, идущей по специфическим законам популяционной генетики, взаимоотношений между ядерными и митохондриальными генетическими системами, необходимо для понимания сложной иерархической организации эвкариотической клетки и организма в целом.
С определенными мутациями в митохондриальной ДНК или в ядерных генах, контролирующих работу митохондрий, связывают некоторые наследственные болезни и старение человека . Накапливаются данные об участии дефектов мтДНК в канцерогенезе. Следовательно, митохондрии могут быть мишенью химиотерапии рака. Имеются факты о тесном взаимодействии ядерного и митохондриального геномов в развитии ряда патологий человека. Множественные делеции мтДНК обнаружены у больных с тяжелой мышечной слабостью, атаксией, глухотой, умственной отсталостью, наследующихся по аутосомно-доминантному типу. Установлен половой диморфизм в клинических проявлениях ишемической болезни сердца, что скорее всего обусловлено материнским эффектом - цитоплазматической наследственностью. Развитие генной терапии внушает надежду на исправление дефектов в геномах митохондрий в обозримом будущем.
Работа выполнена при поддержке Российского
фонда фундаментальных исследований. Проект 01-04-48971.
Автор признателен аспиранту М.К.Иванову,
создавшему рисунки к статье.
Литература
1. Янковский Н.К., Боринская С.А.
Наша история,
записанная в ДНК // Природа. 2001. №6. С.10-18. 2. Минченко А.Г., Дударева Н.А.
Митохондриальный
геном. Новосибирск, 1990. 3. Гвоздев В.А.
// Сорос. образоват. журн. 1999.
№10. С.11-17. 4. Маргелис Л.
Роль симбиоза в эволюции клетки.
М., 1983. 5. Скулачев В.П.
// Сорос. образоват. журн. 1998.
№8. С.2-7. 6. Игамбердиев А.У.
// Сорос. образоват. журн.
2000. №1. С.32-36.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Генетика митохондрий
1. Формальная генетика митохондрий
В отличие от пластид, митохондрии содержатся у всех эукариот: у растений, животных и грибов. Митохондрии всех трех царств выполняют одну и ту же функцию, и структура у них в целом сходна. Митохондрии представляют собой округлые структуры размером от 1 мкм (рис. 1).
Рис. 1 Электронная микрофотография митохондрий листового мезофилла
Однако, в ряде случаев митохондрии могут быть объединены в достаточно длинную трубчатую изогнутую структуру. Внутреннее содержимое митохондрий называется матрикс. Матрикс содержит тонкие фибриллы и гранулы. Было выяснено, что гранулы -- это митохондриальные рибосомы, отличающиеся по размеру и плотности от рибосом цитоплазмы. Митохондрии, как и другие органеллы, окружены наружной двойной мембраной. Наружная мембрана митохондрий сходна с наружной мембраной пластид, ядра и с мембраной эндоплазматического ретикулума. Внутренняя мембрана митохондрий образует впячивания -- кристы. Именно на поверхности внутренней мембраны расположены все основные ферментные ансамбли, обеспечивающие функции митохондрий. Существуют методы разделения внутренней и внешней мембраны митохондрий. Поскольку наружная мембрана митохондрий менее плотная и необратимо набухает в растворе фосфата, это приводит к ее разрыву и отделению от внутренней. После обработки фосфатом изолированных митохондрий при помощи центрифугирования можно разделить внешние и внутренние мембраны этих органелл. Если посмотреть на них в электронный микроскоп, то они выглядят как прозрачные полые сферы, причем объем сферы, образованной внутренней мембраной, намного выше, чем объем сферы внешней мембраны. Поэтому объемную структуру митохондрий легко представить себе как большой шар, помещенный внутри маленького шара. В этом случае у внутренней мембраны возникнут многочисленные складки, так называемые кристы. Активность процессов, протекающих в митохондриях, непосредственно связана с количеством и размером крист. Чем больше поверхность крист и, следовательно, поверхность внутренней мембраны, тем активнее идут эти процессы. Следовательно, внутренняя мембрана митохондрий изменяется в размерах в зависимости от функционального состояния органелл.
Внутренние и наружные мембраны различаются между собой по плотности (внутренняя более плотная), по проницаемости (внутренняя обладает высокоспецифичной проницаемостью, наружная неспецифичной), разным составом ферментов и разным соотношением белков к липидам.
Внутренняя мембрана митохондрий уникальна по своему строению. Она содержит многокомпонентные комплексы белков-ферментов, которые осуществляют перенос электронов, окислительное фосфолирование, синтез цепи жирных кислот, а также белки, которые регулируют перенос небольших молекул во внутреннюю полость митохондрий.
Митохондрии так же, как и пластиды, никогда не возникают «de novo». Даже у организмов, живущих в анаэробных условиях, структуры, подобные митохондриям, есть. Если, например, выращивать один и тот же штамм дрожжей в аэробных и анаэробных условиях, то в клетках, выросших в анаэробных условиях, изменяется размер митохондрий, но не снижается их количество.
Деление митохондрий так же, как и пластид, осуществляется при помощи амитоза, с образованием гантелевидных фигур и их последующей перешнуровкой.
В некоторых случаях удалось показать синхронность деления митохондрий с ядром клетки и достаточно точное их распределение по дочерним клеткам у некоторых биологических объектов. Так, у инфузорий показана полная синхронность деления митохондрий вместе с ядром клетки. В митотически делящихся клетках растений и делящихся сперматоцитах аскариды было показано, что митохондрии достаточно точно распределяются вдоль веретена деления.
Исторически, почти вся формальная генетика митохондрий была изучена на грибах и, главным образом, на дрожжах. У остальных организмов имеются только отдельные факты связи тех или иных признаков с митохондриями. Жизненный цикл дрожжей представлен на рисунке
Рис. 2 Жизненный цикл Saccharomyces cerevisiae
Дрожжи -- одноклеточный, но многоядерный организм. Значительную часть своей жизни они находятся в гаплофазе и, следовательно, их ядра гаплоидны. Гаплоидные клоны, обладающие противоположными половыми факторами (или типами скрещиваемости), А и а, могут сливаться друг с другом. Гаплоидные клоны с одинаковыми типами скрещиваемости не могут участвовать в оплодотворении. После оплодотворения ядра сливаются и образуются диплоидные клоны. В диплоидных клонах происходит споруляция и мейоз, образуется аск, дающий начало гаплоидным клонам двух противоположных типов скрещиваемости А и а в равных пропорциях. Естественно, что простые менделевские гены будут расщепляться точно так же, как и ген, контролирующий половой фактор, т.е. будут давать расщепление 1:1.
Дрожжи в зиготной фазе являются гетерозиготными и могут размножаться двояко: вегетативным и генеративным путем. При вегетативном размножении они просто делятся, и в образовавшиеся клетки попадает несколько диплоидных ядер. Кроме того, вегетативное размножение может происходить и при помощи почкования. В образовавшихся почках ядра тоже, диплоидные. Естественно, что при вегетативном размножении никакого расщепления ядерных генов не происходит -- гетерозиготы остаются гетерозиготами.
При генеративном размножении происходит мейоз и образуются клетки с гаплоидными ядрами, называемые аскоспорами. Аскоспоры гаплоидны, и расщепление у них происходит на равное количество аскоспор с доминантным и рецессивным аллелями, т.е. 1:1.
Таким образом, если расщепления 1:1 не наблюдается, то это могло бы указывать нам на то, что эти гены, возможно, неменделевские и, следовательно, возможно, цитоплазматические.
Существование внеядерного мутанта у дрожжей впервые продемонстрировал французский исследователь Б. Эффрусси еще в 1949 г. Эти мутанты проявляли дефекты дыхания и слабый рост. Они не содержали некоторые цитохромы. Такие мутанты можно было получить в большом количестве (иногда до 100 %) под действием акридиновых красителей. Но и спонтанно они могут возникать с частотой до 1 %. Эти мутанты получили название «petite », от французского слова «маленький».
При скрещивании этих мутантов с нормальными штаммами все потомство было без исключения нормальным. Хотя по другим генетическим маркерам, таким, как потребность в аденине, тиамине, расщепление по факторам полового типа было нормальным -- 1:1.
Если из первого поколения гибридов случайно выбирать клетки и скрещивать их опять с мутантами petite , все потомство было опять нормальным, правда, иногда появлялись и редкие мутантные выщепенцы с частотой менее 1%. Т.е. они появлялись практически с той же частотой, что и спонтанное возникновение этих мутантов. Можно было опять отбирать эти гибриды и скрещивать с нормальными с тем же результатом. Если исходить из того, что это мутации ядерных генов, то это можно было бы представить, как результат расщепления по 20 независимым локусам. Возникновение мутанта с одновременной мутацией в 20 локусах событие практически невероятное.
Р. Райт и Д. Ледерберг получили убедительные доказательства того, что данные мутанты не являются ядерными. Схема их эксперимента была следующей. При слиянии клеток дрожжей ядра сливаются не сразу, и в этот момент можно отсадить почки, содержащие еще гаплоидные ядра как одного, так и другого родителя. Такие гаплоидные почки спонтанно диплоидизируются (А --> АА; а --> аа). Если один штамм, например, с мутацией petite помечен неспособностью расти на аргинине, а второй -- не petite , помечен неспособностью расти на триптофане, то, отбирая почки у таких гибридов, мы отбираем родительские штаммы по ядерным генам. Что же происходит с цитоплазматическими? В результате эксперимента Р. Райта и Д. Ледерберга было выявлено следующее. Из 91 клона было обнаружено 6 клонов, у которых ядро было такое же, как и у не petite мутанта, а фенотип типичный petite . Следовательно, данный фенотип определяется не ядром, а независимо от него, и данную мутацию можно было назвать неядерной.
В дальнейшем были обнаружены и ядерные мутации petite . Всего таких мутантов было обнаружено около 20. Все они нормально менделировали и потомство аскоспор давало нормальное расщепление 2:2, хотя фенотипически они были очень схожи с цитоплазматическими мутантами. При скрещивании цитоплазматических petite с ядерными было обнаружено, что у зигот появляется способность к нормальному дыханию, а затем происходит расщепление 2: Таким образом, тест на комплементарность доказывал то, что мы имеем дело с мутантами разной локализации. Обнаружение ядерных и цитоплазматических мутантов с нарушенной функцией митохондрий указывало также и на то, что не все функции этих органелл кодируются цитоплазматическими генами. Часть из них кодируют ядерные гены.
В дальнейшем Б. Эффрусси обнаружил еще один такой же фенотип, как petite , но наследование данной мутации происходило другим образом. При скрещивании мутантов petite с нормальными клетками все потомство приобретало свойство медленно расти, а расщепление было 0:4. Первый тип цитоплазматических мутантов, который давал только нормальное потомство, в связи с этим был назван нейтральным, а второй, который давал только мутантное, был назван супрессивным, или доминантным, petite . Супрессивность в данном случае это своего рода доминирование. Но это доминирование особого рода, когда рецессивный аллель не просто скрывается в гетерозиготе, он попросту исчезает совсем. Многочисленные опыты показали, что и супрессивные мутанты petite также являются цитоплазматическими, так как факторы, обуславливающие их появление, не наследуются вместе с ядром.
В дальнейшем молекулярные исследования выявили, что супрессивные мутанты petite в отличие от нейтральных обладают более короткими молекулами митохондриальной ДНК, состоящими практически только из АТ-пар. Вероятнее всего, эффект супрессивности основан на более быстром размножении таких митохондриальных ДНК и вследствие этого вытеснением нормальных митохондриальных ДНК.
Таким образом, в цитоплазматических мутантах типа petite имеют место либо сравнительно небольшие делеции в митохондриальной ДНК (нейтральные мутанты petite ), либо тотальные перестройки митохондриального генома -- (супрессивные мутанты petite ).
Кроме того, были обнаружены мутанты с неполной супрессивностью, т.е. способностью давать определенный процент особей нормального типа 10, 20, 30 и даже около 50 процентов.
Оказалось, что степень супрессивности зависит от воздействий внешней среды -- температуры, субстрата и т.д. Ядерные мутанты не проявляли такой зависимости, что позволило отличать неполно супрессивные цитоплазматические petite от ядерных.
После получения данных о цитоплазматических мутантах устойчивости к антибиотикам у хламидомонады, стали получать мутации устойчивости к антибиотикам и у дрожжей. Целый ряд таких мутантов - также оказался цитоплазматическим. При скрещивании, например, эритромицинчувствительных с эритромицинустойчивыми ERs х ERr , все потомство было эритромицинчувствительным Ers (т.е. таким же, как и дикий тип) и не происходило никакого расщепления. Такой же результат был продемонстрирован и с мутантами устойчивости к другим антибиотикам. Однако, если сразу после образования зиготы отбирать почки, то среди них можно найти и мутантные фенотипы.
При дигибридном скрещивании, т.е. при скрещивании двух цитоплазматических мутантов, чувствительных к разным антибиотикам, например устойчивых к хлорамфениколу, но чувствительных к эритромицину с чувствительными к хлорамфениколу, но устойчивыми к эритромицину CrERs х CsERr , в потомстве преобладал фенотип только одного из родителей -- CrERs . Вместе с тем, при отборе из почек сразу после оплодотворения были обнаружены не только родительские классы фенотипов, но и рекомбинанты: CrERr и CsERs , т.е. чувствительные или устойчивые к обоим антибиотикам. Наличие рекомбинантов впервые показало, что и митохондриальные гены могут рекомбинировать так же, как и ядерные. Вместе с тем, в отличие от экспериментов по рекомбинации пластидных генов у хламидомонады, у дрожжей была обнаружена полярность рекомбинации, т.е. неравное количество рекомбинантных фенотипов в зависимости от направления скрещивания. Полярность рекомбинации была объяснена как наличие специального генетического фактора пола в геноме митохондрий. Этот фактор был обозначен как щ+ и щ-. Родительская форма, имеющий фактор щ+, т.е. женский родитель, обеспечивает преимущественную передачу (более высокую частоту передачи) своих маркеров. При скрещивании однополых по этому митохондриальному фактору родителей полярности рекомбинации не наблюдается и получается равное число рекомбинантов. Сам половой фактор митохондрий наследуется независимо от пола организма.
В действительности имеется ли пол у органелл цитоплазмы -- митохондрий в общепринятом смысле? Можно считать, что имеется, если мы считаем, что у кишечной палочки он есть.
Но главное заключалось в том, что при помощи множества полученных мутаций и обнаружения рекомбинации митохондриальных генов стало возможным их картирование.
В экспериментах по скрещиванию мутаций типа petite с мутациями устойчивости к антибиотикам было выяснено, что, по крайней мере, все супрессивные мутации petite в скрещиваниях утрачивают гены устойчивости к антибиотикам. Как было выяснено, это происходит потому, что супрессивные petite имеют обширные области поражения митохондриальной ДНК, и в этом случае ожидать рекомбинации просто невозможно. При индукции мутаций дыхательной недостаточности у мутантов с устойчивостью к тем или иным антибиотикам оказывалось, что иногда маркеры устойчивости утрачивались. При получении мутантов с дыхательной недостаточностью, используя в качестве исходной формы мутантов с двойной устойчивостью к антибиотикам, у полученных дефектных по дыханию мутантов могли быть потеряны оба маркера устойчивости или только один из них. Это свидетельствовало о том, что мутанты с дыхательной недостаточностью представляют собой ту или иную степень делетирования митохондриальной ДНК, и, следовательно, это также можно было использовать для картирования генома митохондрий.
У нейроспоры в 1952 г. К. Митчелом был обнаружен первый медленно растущий мутант, названный впоследствии MI -1 (сокращение от английского «материнское наследование» -- maternal inheritance ). Наследование этой мутации происходило в зависимости от направления скрещивания, и все потомство было таким же по фенотипу, как и материнская форма. Вероятно, это происходит потому, что при оплодотворении мужская гамета у нейроспоры не привносит цитоплазмы. На связь этой спонтанно возникшей мутации с митохондриями указывало не только материнское наследование и различия в реципрокных скрещиваниях, но и то, что у них отсутствовали цитохромы a и b в системе переноса электронов.
Впоследствии были получены и другие медленно растущие штаммы у нейроспоры, связанные с дыхательной недостаточностью митохондрий. Некоторые из них, например, мутанты MI -3 и MI -4, как оказалось, наследовались так же, как и мутант MI -1, в то время как другая часть, например, мутанты С115 и С117 проявляли обычное менделевское моногибридное наследование. Это напоминает и о других аналогичных случаях, когда фенотип органелл, и хлоропластов, и митохондрий изменяется при возникновении как ядерных, так и цитоплазматических мутаций, что свидетельствует, что и цитоплазматические и ядерные генетические системы совместно контролируют их функции.
В дальнейшем было обнаружено несколько генов супрессоров, введение которых восстанавливало скорость роста у медленно растущих мутантов. Интересно отметить, что каждый из этих супрессоров восстанавливал скорость роста только у одного из мутантов. Например, ген супрессор, названный f , восстанавливал скорость роста у цитоплазматичкого мутанта MI -1, но не у другого цитоплазматического мутанта MI -3 или MI -4, и не у ядерных мутантов С115 и С117 . Аналогичным образом действовали и другие супрессоры. Если через множество поколений путем скрещиваний вывести гены супрессоры из грибов, то мутантный цитоплазматический фенотип снова проявится. Аналогичное взаимодействие ядерных и цитоплазматических генов можно наблюдать и у высших растений, например, при наследовании признака мужской стерильности у многих растений.
При скрещивании ядерных и цитоплазматических медленно растущих мутантов между собой было показано независимое наследование ядерных и цитоплазматических генов.
Например, при скрещивании дикий тип х (MI -1 х С115 ) потомство F 1 (MI -1 х С115 ) было фенотипически однородным -- все особи были медленно растущими, а потомство возвратных или анализирующих скрещиваний дикий тип х (MI -1 х С115 ) уже не содержало мутаций MI -1 и расщеплялось по ядерному гену С-115 в соотношении 1:1.
Скрещивание цитоплазматических мутантов между собой не давало каких-то новых результатов, так как цитоплазматические мутанты, по крайней мере, у нейроспоры при половом воспроизведении демонстрируют строго материнское наследование. Между тем разные цитоплазматические мутанты, хотя и обладали в принципе одинаковым фенотипом -- замедленным ростом -- фенотипические различия между ними все же можно было выявлять, так как они обладали разной степенью замедления этого роста. Однако строгое материнское наследование при половом воспроизведении не позволяло объединить в цитогету (цитоплазматическую гетерозиготу) две цитоплазматические мутации, что делало невозможным рекомбинации цитоплазматических генов и, следовательно, их картирование.
Выход из этого положения был найден при помощи слияния гифов нейроспоры, что позволило соединять в одной клетке различные как ядерные, так и неядерные геномы.
При создании различных цитогет были получены следующие результаты:
MI -1 / дикий тип -- все потомство только дикого типа;
MI -3 / дикий тип -- часть потомства дикого типа, а другая часть растет со скоростью, свойственной мутанту MI -3;
MI -1 / MI -З -- большая часть потомства с фенотипом MI -3 и небольшая часть потомства с фенотипом MI -1;
MI -1 / MI -4 -- первоначально фенотип, свойственный дикому типу, а затем расщепление на фенотипы MI -1 и MI -4.
Таким образом, в последнем случае была обнаружена комплементация цитоплазматических мутаций, что свидетельствует, что эти мутации произошли в разных участках митохондриального генома.
В дальнейшем были получены и другие цитоплазматические мутации нейроспоры. Метод слияния гиф и получения при этом цитогет позволял надеяться на получение различных рекомбинантов и последующего построения генетической карты нейроспоры. Однако этому воспрепятствовало то обстоятельство, что у нейроспоры не было получено большого разнообразия цитоплазматических мутаций такого, как у хламидомонады или дрожжей.
Впоследствии различные нехромосомные мутации, полученные у нейроспоры, исследовали при помощи методов молекулярной биологии, и их удалось связать с митохондриальным геномом.
У другого гриба подоспоры была обнаружена мутация, вызывающая феномен преждевременного старения. У мутантов постепенно снижалась жизнеспособность культуры при пересеве. При реципрокных скрещиваниях был выяснен материнский характер наследования феномена старения. Вместе с тем, материнское наследование было неполным. Передача признака осуществляется как половым путем, так и путем соединения мицелиев. Наличие расщепления, хотя и нерегулярного, указывает на корпускулярную природу наследования признака. Было проведено достаточно много исследований, которые позволили показать, что это неинфекционный агент, а митохондриальный ген. Хотя в настоящее время нет полных молекулярных данных, уже ясно, что это также мутации митохондриального генома. Наличие гена старения в митохондриальном геноме породило массу спекуляций на геронтологические темы, и некоторые медики считают, что старение и у человека связано не только с изменением функций митохондрий, но и с изменением их генома.
Несмотря на спекулятивность идеи о связи геронтологических процессов у человека с изменениями митохондриальной ДНК, новые данные по изучению изменчивости генома митохондрий человека это подтверждают.
Издавна у человека было известно достаточно большое количество заболеваний, наследующихся по материнской линии -- от матери ко всем потомкам. Эти болезни достаточно редко распространены, вероятно, и в силу того, что передаются только женским полом. Кроме того, большие делеционные изменения в митохондриальной ДНК, конечно, чаще всего приводят либо к летальному исходу еще в эмбриональном периоде, либо к нарушению репродукционных функций. В любом случае они эффективно отметаются естественным отбором.
Формальный генетический подход, который был достаточно хорошо применен для исследования цитоплазматических генов у модельных объектов (хламидомонада, дрожжи и др.), не был столь успешным для анализа цитоплазматически наследуемых признаков у человека и поэтому самое большее, что удалось выяснить из анализа родословных, это то, что такие наследственные болезни все же существуют.
Кроме хорошо известного синдрома -- атрофии зрительного нерва (болезнь Лебера или наследственная оптическая невропатия) существуют и другие болезни, наследующиеся по внеядерному типу. Эти болезни связаны, прежде всего, с нарушением функционирования мышц, работы мозга, сердца, эндокринных систем и связаны с недостаточно активной функцией митохондрий в тех или других органах. Существует даже митохондриально обусловленная форма диабета.
Только с помощью молекулярных методов удалось выявить природу этих болезней. Исследование различных семей с болезнью Лебера показало, что в разных случаях имеют место мутации в различных участках митохондриального генома.
Чаще всего семьи с наследственными цитоплазматическими болезнями проявляют гетероплазмию и матери имеют как нормальные, так и мутантные митохондриальные ДНК, в результате чего выщепляются потомки как с мутантным, так и нормальным плазматипом.
Связь между возрастом человека и митохондриальной ДНК также была показана при помощи методов молекулярной биологии. Исследования митохондриальной ДНК у людей различного возраста показали, что у пожилых людей быстро увеличивается процент мутантной митохондриальной ДНК в клетках мозга и сердца. Кроме того, исследования некоторых наследственных синдромов показывают, что обладающие ими пациенты имеют и повышенную частоту мутаций митохондриальной ДНК, что возможно и является причиной сокращения продолжительности жизни.
Кроме мутаций митохондриального генома, приводящих к серьезным патологиям организма, было обнаружено множество, в достаточной степени, нейтральных мутаций митохондриального генома среди различных популяций рас человека. Эти обширные исследования тысяч людей со всех континентов помогают восстанавливать происхождение и эволюцию человека. Сравнивая митохондриальную ДНК человека с ДНК человекообразных обезьян (горилла, орангутанг, шимпанзе) и исходя из того, что дивергенция человека и человекообразных приматов произошла примерно 13 миллионов лет назад, можно рассчитать количество лет, необходимых для изменения одной пары оснований. В дальнейшем, сравнивая дивергенцию митохондриальной ДНК у различных человеческих рас, можно было определить место рождения первой женщины, можно сказать Евы, и время расселения человека по различным континентам (рис. 3).
Размещено на http://www.allbest.ru/
Рис. 3 Расселение человека, согласно данным Д. Уоллеса, по анализу изменчивости митохондриальных ДНК. Цифрами обозначено время заселения данной территории в тысячах лет тому назад
Поскольку самая изменчивая митохондриальная ДНК была обнаружены среди аборигенов Африки, то можно предположить, что «праматерью» человеческого рода была африканская женщина. Произошло это приблизительно 100 000 лет тому назад. Приблизительно 70 000 лет тому назад человек начал заселять центральную Азию через Ближний Восток и Саудовскую Аравию, чуть позже Юго-Восточную Азию, Индонезию и Австралию. Около 50 000 лет тому назад люди появились в Европе. Эти же данные показали, что заселение Американского континента происходило в два этапа: сначала 30 000 лет тому назад через Беренгию (существовавшую в то время сушу, связывающую Америку и Азию) с Севера до самого юга Американского континента, а затем 8 000 лет тому назад также из Северо-Восточной Азии на восток Северной Америки. Переселенцы на островах Тихого океана появились сравнительно недавно -- несколько тысяч лет тому назад.
Следует отметить, что эти данные, основанные на сравнительном анализе митохондриальной ДНК, достаточно хорошо согласуются как с археологическими данными, так и с данными лингвистического анализа.
Использование именно митохондриальной ДНК для анализа истории человечества стало возможным потому, что митохондриальный геном имеет сравнительно небольшой размер, наследуется исключительно по материнской линии и в отличие от ядерных генов не рекомбинирует.
Геном митохондрий
Митохондрии содержатся не только в клетках растений, но также и в клетках животных и грибов. Эти органеллы более универсальны, чем пластиды. Впервые ДНК в митохондриях были открыты в 1963 году (М. Наас) сразу же после открытия ДНК в пластидах. Несмотря на сходство функций и структуры митохондрий всех трех царств эукариот, их генетическая организация достаточно сильно различается, поэтому обычно организацию геномов митохондрий у этих царств рассматривают отдельно, выявляя при этом общие черты организации генома.
Физико-химический состав ДНК митохондрий у различных царств различен. У растений он довольно постоянен: от 45 до 47 % ДНК состоит из ГЦ-пар. У животных и грибов -- варьирует более значительно: от 21 до 50 % ГЦ-пар.
У многоклеточных животных размеры генома митохондрий колеблются от 14.5 до 19.5 т.п.н. Практически, это всегда одна кольцевая молекула ДНК. Например, ДНК митохондрий человека -- кольцевая молекула размером 16 569 пар нуклеотидов. Этот размер можно выразить и в других единицах -- в виде молекулярной массы -- 10 6 дальтон или в виде длины контура молекулы -- 5 мкм. Первичная структура этой молекулы полностью определена. В митохондриях содержится собственный аппарат трансляции -- т.е. собственные рибосомы 70S, похожие на хлоропластные или прокариотические и состоящие из двух субъединиц, собственные матричные РНК, необходимые ферменты и белковые факторы. В их геноме закодированы 12S- и 16S - рибосомальные РНК, а так же 22 транспортные РНК. Кроме того, митохондриальная ДНК кодирует 13 полипептидов, из которых 12 идентифицированы. Все кодирующие последовательности расположены прямо друг за другом. В крайнем случае, они разделены лишь несколькими нуклеотидами. Некодирующие последовательности, т.е. интроны отсутствуют. Вслед за кодирующей последовательностью почти всегда находится ген транспортной РНК. Например, порядок таков: транспортная РНК фенилаланина -- ген 12S рибосомальной РНК -- транспортная РНК валина -- ген 16S рибосомальной РНК -- транспортная РНК лейцина и т.д. Такой порядок характерен не только для митохондрий человека, он очень консервативен и характерен для всех животных: дрозофилы, быка, мыши, птиц, рептилий и др. животных.
Большая часть генов расположена в тяжелой цепи, в легкой цепи только гены восьми транспортных РНК и один структурный ген. Таким образом, в отличие от всех других геномов, в геноме митохондрий обе цепи смысловые.
Хотя порядок генов у митохондрий животных и одинаков, выяснено, что сами гены обладают различной консервативностью. Наиболее вариабельна последовательность нуклеотидов участка начала репликации и ряд структурных генов. Наиболее консервативные последовательности расположены в генах рибосомальных РНК и некоторых структурных генах, в том числе в кодирующей последовательности АТФ-азы.
Следует отметить, что универсальность генетического кода нарушена в геноме митохондрий. Например, митохондрии человека используют триплет AUA в качестве кодона для метионина, а не для изолейцина, как у всех, а триплет UGA, используемый в стандартном генетическом словаре как терминирующий кодон, у митохондрий кодирует триптофан.
В целом митохондриальная ДНК человека выглядит так же, как и других млекопитающих: мыши и быка. Несмотря на то, что это далеко не близкие виды -- размеры их митохондриальных ДНК довольно близки между собой: 16 569; 16 295; и 16 338 пар оснований, соответственно. Гены транспортной РНК разделяют некоторые смысловые гены. Наиболее важные из структурных генов -- гены цитохромоксидазы, NADH-дегидрогеназы, цитохром-С оксидоредуктазы и АТФ-синтетазы (рис. 4).
На карте митохондриального генома человека, кроме генов показано и пять хорошо известных болезней человека, наследующихся по материнской линии и вызванных мутациями в митохондриальном геноме.
Так, например, болезнь Лебера -- атрофия зрительного нерва -- вызвана мутацией в гене NADH дегидрогеназы. Эта же болезнь может быть вызвана и мутацией в гене цитохрома b и других локусов. Всего известно нарушение четырех локусов, способных вызвать тот же мутантный фенотип. Кроме того, на этой же карте показано еще четыре болезни, связанные с дефектами мозга, мышц, сердца, почек и печени. Все эти болезни наследуются по материнской линии, и если мать имеет не только дефектные, но и нормальные митохондриальные ДНК и митохондрии, то, происходит сортировка мутантных и нормальных органелл, и потомство может иметь и те, и другие органеллы в различных пропорциях, и мы можем наблюдать также и соматическое расщепление, когда отдельные части тела не будут иметь этих дефектов.
Размещено на http://www.allbest.ru/
Рис. 4 Структура митохондриального генома млекопитающих, основанная на полном сиквенсе митохондриальной ДНК человека, мыши и быка
Таким образом, небольшой по размерам митохондриальный геном животных может кодировать чрезвычайно важные функции организма и в значительной степени определять его нормальное развитие.
Так же, как и геном пластид, геном митохондрий кодирует только часть митохондриальных полипептидов (табл. 1) и наблюдается феномен двойного кодирования. Например, часть субъединиц АТФ-азного комплекса кодируется ядром, в то время как другая часть -- геномом митохондрий. Большая часть генов, кодирующих рибосомальные миохондриальные РНК и белки, а также ферменты транскрипции и трансляции, кодируется ядром клетки.
Таблица 1
Гены митохондриальных ДНК животных
митохондрия геном нейроспора мезофилл
генома животных:
1. компактное расположение генов на мтДНК;
отсутствие в генах интронов;
3. отсутствие некодирующих участков в мтДНК, кроме областей ORI;
4. расположение генов тРНК между другими генами;
5. высокое сходство в размерах генома и в расположении генов у различных видов;
6. наличие одного ORI для каждой нити мтДНК;
7. симметричная транскрипция обеих нитей;
8. наличие одной, в принципе, области инициации транскрипции для каждой нити ДНК;
9. отсутствие 5 / - и 3 / - концевых некодирующих последовательностей в мРНК;
10. созревание мРНК в результате расщепления первичного транскрипта по последовательностям тРНК.
У грибов размеры генома митохондрий в среднем значительно больше и колеблются от 17,3 до 101 т.п.н. Причем в дополнении к основной, как правило, кольцевой молекуле ДНК обнаруживают и от одной до 4-х плазмидоподобных кольцевых или линейных молекул размером от 1 до 13 т.п.н. Размеры митохондриального генома у дрожжей варьируют не только между различными видами, но и даже между различными штаммами. Основные причины существенных различий в геноме митохондрий у грибов заключаются в наличии - отсутствии интронов. У разных видов дрожжей, например, размер митохондриальной ДНК колеблется от 57 до 85 т.п.н.
Наличие интронов и молекул митохондриальной ДНК различного размерного класса является наиболее характерной чертой, отличающей митохондрии грибов от митохондрий животных. Интроны разрывают многие последовательности -- гены рибосомальной РНК, гены некоторых структурных белков, кодирующих митохондриальные ферменты. Наличие большинства интронов не является обязательным для нормального функционирования митохондрий. Искусственно сконструированы штаммы дрожжей, полностью лишенные митохондриальных интронов.
Многие интроны митохондриальной ДНК дрожжей содержат открытые рамки считывания, которые кодируют мутуразы, участвующие в сплайсинге, тогда как другие интроны содержат кодирующие последовательности эндонуклеаз и даже обратных транскиптаз.
Все гены, обнаруженные в митохондриальной ДНК животных, присутствуют и у грибов. Кроме того, у грибов обнаружены и другие гены: у них большее число генов тРНК, обнаружены гены 6-й, 8-й и 9-й субъединиц АТФ-азного комплекса, ряд новых структурных генов и ряд генов с неизвестной функцией (табл. 2).
Таблица 2
Гены митохондриальных ДНК дрожжей
|
Компоненты митохондрий |
||
|
Рибосомальные РНК |
rns(21 S), rnl(1 5 S) |
|
|
Рибосомальные белки: малая субъединица |
||
|
Транспортные РНК |
||
|
Цитохром b (комплекс III) |
с ob (или cyb ) |
|
|
Цитохром с оксидаза (комплекс IV) |
cox 1, cox 2, coxd 3 |
|
|
АТФ-синтаза |
atp6, atp8, atp9 |
|
|
Интрон-кодируемые off: РНК-матуразы Эндонуклеазы Белки, подобные обратной трансриптазе |
aI1, aI2 |
|
|
Неидентифицированные рамки считывания |
В митохондриальной ДНК дрожжей обнаружено только 2 гена рибосомальной РНК и только 1 ген рибосомальных белков. Этот белок расположен в малой субъединице рибосомы. Ген рибосомального белка достаточно вариабелен по размерам даже у разных штаммов, за что и получил название вариабельного (Var l). Остальные белки и РНК митохондриальных рибосом кодируются ядерными генами. 24 гена транспортной РНК обеспечивают транспорт всех аминокислот к месту синтеза белка и только одна транспортная РНК, транспортирующая лизин, импортируется из цитоплазмы и кодируется ядром. Все транспортные РНК митохондрий дрожжей кодируются одной и той же нитью ДНК и только одна из них -- противоположной нитью. Ни один из генов транспортной ДНК не имеет интронов. Гены белков цитохрома b и гены белков цитохрома С могут иметь множество интронов -- от 5 до 9.
Из приведенных данных следует, что кодируемых митохондриальным геномом дрожжей структурных белков явно недостаточно для функционирования этих органелл и большая часть из них кодируется ядерным геномом.
Характерные черты организации и экспрессии митохондриального генома грибов:
1. значительное разнообразие в наборах и расположении митохондриальных генов у различных видов;
большое разнообразие способов организации генетического материала - от компактной организации генома до свободного распределения генов по мтДНК при протяжённых некодирующих последовательностях между генами;
3. мозаичное строение ряда генов;
4. значительные внутривидовые вариации размеров мтДНК, связанные с наличием "необязательных" интронов;
5. способность отдельных сегментов мтДНК к выщеплению и амплификации с образованием дефектного митохондриального генома;
6. наличие одного или нескольких ORI, в каждом из которых репликация инициируется двунаправлено;
7. расположение всех митохондриальных генов на одной нити мтДНК и асимметричная транскрипция мтДНК;
8.множественность транскрипционных единиц мтДНК;
9. разнообразие сигналов процессинга первичных транскриптов, в качестве которых могут использоваться и тРНК, и олигонуклеотидные блоки другого типа - в зависимости от вида;
10. в большинстве случаев мРНК содержат протяжённые концевые некодирующие последовательности.
Наиболее сложная организация митохондриального генома у высших растений. У них митохондриальный геном представляет собой набор суперскрученных двуцепочечных кольцевых и/или линейных молекул. Все последовательности митохондриального генома могут быть организованы в одну крупную кольцевую «хромосому», а наблюдаемые различные размерные классы митохондриальных ДНК, скорее всего, являются результатом рекомбинационных процессов. По крайней мере, на шпинате, видах двух родов Brassica и Raphanus , сахарной свекле и пшенице было показано, что причина такой диспергированности митохондриального генома заключается в рекомбинации гомологичных участков митохондриальной ДНК. Благодаря наличию прямо ориентированных двух-трех семейств повторов размером от 1 до 14 т.п.н., молекулы митохондриальной ДНК способны к активным меж- и внутригеномным перестройкам. В результате таких перестроек митохондриальная ДНК может присутствовать в виде молекул различного размерного класса.
Так, например, у крестоцветного Brassica campestris митохондриальная ДНК присутствует в виде кольцевых молекул трех типов. Первый тип содержит полный геном -- 218 т.п.н., второй - 135 и третий -- 83 т.п.н. Субгеномные кольца образуются в результате рекомбинации геномных колец, имеющих пару прямых повторов длиной 2 т.п.н.
У пшеницы размер митохондриального генома значительно больше -- 430 т.п.н., и имеется более 10 прямых рекомбинационных повторов, в результате при электронно-микроскопическом наблюдении удается увидеть множество колец различного размера, но никто не наблюдал одну большую кольцевую молекулу, возможно, в этом состоянии митохондриальный геном пшеницы никогда и не присутствует. У мха маршанции и другого крестоцветного Brassica hirta прямые рекомбинационные повторы отсутствуют и, возможно, именно поэтому митохондриальная ДНК находиться в виде кольцевых молекул одного размерного класса. Однако для митохондриальной ДНК высших растений это скорее исключение, чем правило. У большинства высших растений в митохондриальном геноме присутствуют как рекомбинационные повторы, так и молекулы митохондриальной ДНК различного размерного класса.
Количество молекул одного размерного класса может различаться очень значительно в различных тканях растения, в зависимости от состояния растений и условий окружающей среды. Отмечено изменение численных соотношений молекул митохондриальной ДНК разного размерного класса при культивировании растений in vivo и in vitro . Возможно, изменение численных отношений между молекулами различных размерных классов отражает адаптивность растений путем повышенной амплификации нужных генов.
Кроме того, в митохондриальной геноме могут присутствовать и плазмиды как линейные, так и кольцевые, как с ДНК-ми, так и РНК-ми последовательностями, размером от 1 до 30 т.п.н. Митохондриальные плазмиды, вероятно, произошли от других клеточных геномов или даже других организмов. Иногда их наличие или отсутствие удается связать с цитоплазматической мужской стерильностью растений, но, правда, не всегда. Плазмиды у некоторых видов присутствуют, а стерильности не наблюдается. По крайней мере, в одном случае достаточно четко продемонстрировано, что в митохондриях линий с так называемым S-типом стерильности кукурузы обнаружена корреляция между присутствием плазмидоподобной митохондриальной ДНК и проявлением феномена цитоплазматической мужской стерильности. Была отмечена способность митохондриальных плазмид встраиваться, как в геном митохондрий, так и в хромосомы ядра. Однако, в других случаях присутствие плазмидной ДНК не всегда вызывает стерильность пыльцы.
Размер митохондриального генома растений наиболее вариабелен -- от 200 до 2500 т.п.н. Размер митохондриального генома высших растений больше, чем размер их хлоропластного генома.
Значительное варьирование размеров митохондриального генома является второй особенностью митохондриального генома растений. Геном не только очень велик, но и может быть различным, даже у близкородственных видов, причем в одних случаях может наблюдаться низкая вариабельность -- виды рода Brassica , в других -- очень большая. Самая высокая вариабельность размеров наблюдается у тыквенных. В пределах этого семейства размер митохондриального генома наиболее изменчив -- от 330 т.п.н. у арбуза до 2500 т.п.н. у дыни. Поэтому доля митохондриальной ДНК в общем объеме генома растений также может значительно изменяться -- порядка 1 % у большинства растений, до 15 % в клетках гипокотиля дыни.
Наличие у митохондриальных геномов больших размеров пытаются объяснить различными причинами.
Наличие дополнительных генов или особых последовательностей, необходимых для функционирования митохондрий.
Присутствие ДНК, которая используется растением, но не в качестве кодирующей, а какой-то другой функции.
ДНК, которая не используется для функционирования митохондрий, так называемая «эгоистичная» ДНК.
По-видимому, существует еще одна возможность для увеличения размеров митохондриального генома -- это последовательности, гомологичные ядерной и хлоропластной ДНК. Последовательности, гомологичные ядерной ДНК, например, у арабидопсис составляют до 5 % митохондриального генома. Первоначально последовательность хлоропластного генома, инкорпорированная в митохондриальный, была обнаружена у кукурузы. Она включала участок около 14 т.п.н., содержащий измененные хлоропластные гены 16S-рибосомальной РНК и участок большой субъединицы РДФК/О. Впоследствии хлоропластные вставки были обнаружены в митохондриальном геноме многих видов высших растений. Обычно, они составляют 1 -- 2 % от митохондриальных последовательностей и включают три главные последовательности.
Последовательность длиной 12 т.п.о. из обратного повтора хлоропластной ДНК. Она содержит последовательности для 3" экзона четырех транспортных РНК и последовательность 16S рибосомальной РНК.
Последовательность длиной от 1,9 до 2,7 т.п.н., которая полностью кодирует большую субъединицу РБФК/О.
Последовательность длиной не более 2 т.п.о. В хлоропластом геноме этот участок кодирует 3" -- конец 23S-pибоcoмальной РНК, 4,5S- и 5S-pPHK, а также три транспортные РНК. Из всех последовательностей хлоропластного генома, которые присутствуют в митохондриальном геноме растений, только последовательности транспортной РНК действительно транскрибируются.
Поскольку у многих видов растений в митохондриальном геноме присутствуют одни и те же хлоропластные последовательности, можно предположить, что они имеют какое-либо функциональное значение. В то же время их роль, механизм переноса и время этого переноса остаются неизвестными. Произошел ли этот перенос в отдаленное в эволюции время становления эукариотической клетки или же наличие хлоропластных вставок в митохондриальном геноме свидетельствует о том, что это нормальный процесс обмена информации между органеллами, происходящий и сейчас, либо он происходит периодически в относительно недавнем эволюционном времени становления конкретных видов и родов растений?
Кроме того, часть последовательностей митохондриального генома представляет собой последовательности, гомологичные вирусным.
Для установления числа генов в геноме митохондрий растений, которые действительно функционируют, рядом исследователей было определено количество продуктов трансляции. Было показано, что число обнаруживаемых белковых полос было одинаковым даже для растений с 10-кратными различиями по величине генома. Хотя использованные методы и не дают прямого ответа на вопрос об общем числе генов митохондриального генома, тем не менее, интересно, что одно и то же число продуктов трансляции было выявлено у анализируемых видов покрытосеменных и было близко к числу генов, кодирующих белки у митохондрий животных и дрожжей.
Впервые полная нуклеотидная последовательность митохондриальной ДНК у растений была определена в 1986 г. у одного вида -- маршанции (Marchantia polymorpha ), а позже у арабидопсиса и у нескольких видов водорослей.
Молекула митохондриальной ДНК у маршанции имеет размер 186 608 п.н. Она кодирует гены 3 рРНК, 29 генов для 27 тРНК и 30 генов известных функциональных белков (16 рибосомальных белков, 3 субъединицы цитохром С оксидазы, цитохрома b, 4 субъединицы АТФ-синтетазы и 9 субъединиц НАДН-дегидрогеназы). Геном также содержит 32 неидентифицированные открытые рамки считывания. Кроме того, обнаружено 32 интрона, расположенных в 16 генах. У разных растений количество генов того или иного комплекса может варьировать, поскольку один или более генов этого комплекса могут быть перенесены в ядро. Среди неидентифицированных генов, по крайней мере, 10 постоянно встречаются практически у всех видов растений, что свидетельствует о важности их функций.
Число митохондриальных генов, кодирующих транспортные РНК митохондрий растений, в значительной степени изменчиво. У многих видов собственных митохондриальных транспортных РНК явно недостаточно, и поэтому они экспортируются из цитоплазмы (кодируются ядром или пластидным геномом). Так, например, у арабидопсиса 12 транспортных РНК имеют митохондриальное кодирование, 6 -- хлоропластное и 13 -- ядерное; у маршанции 29 -- митохондриальное и 2 -- ядерное, и ни одна из транспортных РНК не имеет хлоропластного кодирования; у картофеля 25 -- митохондриальное, 5 -- хлоропластное и 11 -- ядерное; у пшеницы, 9 -- митохондриальное, 6 -- хлоропластное и 3 -- ядерное (табл. 3).
В отличие от митохондриальной ДНК животных и хлоропластных генов, гены митохондриальной ДНК растений дисперсно распределены в геноме. Это касается как генов, кодирующих транспортные РНК, так и генов, кодирующих белки.
Таблица 3
Природа митохондриальных транспортных РНК у растений
|
Количество транспортных РНК, кодируемых геномами |
||||
|
органелл |
||||
|
митохондрий |
хлоропластов |
|||
|
Арабидопсис |
||||
|
Маршанция |
||||
|
Картофель |
||||
|
Не определено |
Не определено |
|||
|
Подсолнечник |
Не определено |
|||
|
Не определено |
||||
|
Кукуруза |
Не определено |
Как и геном митохондрий грибов, геном митохондрий растений имеет интроны, которых не имеют геномы митохондрий животных.
У некоторых видов ряд генов в геноме продублирован. Так, у кукурузы и кормовых бобов гены рРНК не повторены, а у пшеницы они повторены несколько раз. Гены, кодирующие белки митохондрий, также могут быть повторены в их геноме.
Естественно, что митохондрии так же, как и хлоропласты, содержат гораздо больше белков-ферментов, чем их геном генов. И, следовательно, большинство белков контролируются ядерным геномом, собираются в цитоплазме на цитоплазматических, а не на митохондриальных рибосомах и транспортируются в митохондриальные мембраны.
Таким образом, митохондриальный геном растений представляет собой крайне изменчивую по структуре, но достаточно стабильную по числу генов систему. В отличие от компактного генома хлоропластов, в митохондриальном геноме растений гены составляют менее 20 % генома. Увеличение генома митохондрий по сравнению с грибами или животными вызвано наличием интронов, различных повторяющихся последовательностей, вставок из генома хлоропластов, ядра и вирусов. Функции примерно 50 % митохондриального генома растений пока не выяснены. Кроме того, что многие структурные гены, контролирующие функцию митохондрий, находятся в ядре, там же находятся и многие гены, контролирующие процессы транскрипции, процессинга, трансляции митохондриальных генов. Следовательно, митохондрии являются даже менее автономными органеллами, чем пластиды.
Литература
Основная:
1. Алехина Н.Д., Балнокин Ю.В., Гавриленко В.Ф. и др. Физиология растений. Учебник для студ. Вузов. М.: Академия. 2005. 640 с.
Давыденко О.Г. Нехромосомная наследственность. Минск: БГУ. 2001. 189 с.
3. Даниленко Н.Г., Давыденко О.Г. Миры геномов органелл. Минск: Техналогия. 2003. 494 с.
4. Иванов В.И. и др. Генетика. М.: Академкнига. 2006. 638 с.
5. Жимулев И.С. Общая и молекулярная генетика. Новосибирск: Сиб. унив. 2007. 479 с.
6. Сингер М., Берг П. Гены и геномы. М.: Мир. 1998. Т. 1-
7. Ченцов Ю. С. Введение в клеточную биологию. М.: Академкнига. 2004. 495 с.
Дополнительная:
1. Даниленко Н.Г. РНК-редактирование: генетическая информация корректируется после транскрипции // Генетика. 2001. Т. 37. №3. С. 294-316.
Маргелис Л. Роль симбиоза в эволюции клетки. М.: Мир, 1983.
3. Одинцова М. С., Юрина Н. П. Геном митохондрий протистов // Генетика. 200 Т. 38. №6. С. 773-778.
4. Одинцова М. С., Юрина Н. П. Геном пластид высших растений и водорослей: структура и функции // Мол. Биол. 2003. Т. 37. № 5. С. 768-783.
5. Юрина Н. П., Одинцова М. С. Общие черты организации генома хлоропластов. Сравнение с геномами про- и эукариот // Мол. Биол. 199 Т. 36. № 4. С. 757-771.
6. Юрина Н. П., Одинцова М. С. Сравнительная характеристика структурной организации геномов хлоропластов и митохондрий растений// Генетика. 1998. Т. 34. №1. С. 5-2.
Размещено на Allbest.ru
...Подобные документы
Сущность ультраструктурной организации митохондрий. Роль митохондрий в поддержании окислительно-восстановительного баланса клетки. Специфика энергетических функций митохондрий. Изменение морфофункциональных характеристик митохондрий при ацидозе.
дипломная работа , добавлен 27.01.2018
Исследование функциональной роли и структурной организации митохондрий. Рассмотрение и характеристика работы дыхательной цепи митохондрий в условиях нормоксии. Ознакомление с антигипоксическим действием нейротрофического фактора головного мозга.
курсовая работа , добавлен 18.04.2018
Основные механизмы клеточной гибели. Митохондрия как центральный контрольный пункт апоптоза. Морфологические изменения и перераспределение митохондрий в клетке во время апоптоза. Модели высвобождения цитохрома С. Роль митохондрий в процессе старения.
курсовая работа , добавлен 07.01.2013
Комплекс ферментов, локализованных на внутренней мембране митохондрий. Процесс окислительного фосфорилирования. Синтез АТФ на внутренней мембране митохондрий в присутствии кислорода. Компоненты дыхательной цепи. Суть хемиосмотической теории П. Митчелла.
презентация , добавлен 22.10.2014
Изучение плана строения митохондрий и пластид, их функций. Гипотеза о симбиотическом происхождении митохондрий и хлоропластов. Общая типовая характеристика мышечной ткани. Сперматогенез, его основные периоды: размножение, рост, созревание и формирование.
контрольная работа , добавлен 11.03.2014
Понятие и свойства митохондрий, их строение, участие в клеточном дыхании и обмене энергией. Характерные особенности гаструляции эмбрионального развития. Рассмотрение функций, строения, классификации лейкоцитов. Внешний вид тимуса (вилочковой железы).
контрольная работа , добавлен 21.04.2015
Строение, химический состав, распространение в природе и значение таксономической группы слизевиков. Вегетативные тела слизевиков. Трофическая и расселительная стадии. Процесс образования спор. Присутствие в циклах подвижных стадий, строение митохондрий.
курсовая работа , добавлен 12.08.2015
Строение и основные компоненты клеточной оболочки водорослей. Случаи беспорядочного расположения фибрилл среди зеленых водорослей, организация цитоплазмы у различных у различных представителей вида, назначение жгутиков, митохондрий и хлоропластов.
курсовая работа , добавлен 29.07.2009
Клиническое применение фотодинамической терапии. Механизм действия фотосенсибилизаторов на клеточном уровне. Роль митохондрий и ионов кальция в фотодинамически индуцированном апоптозе. Участие сигнальных процессов и защитных белков в реакциях клеток.
контрольная работа , добавлен 19.08.2015
Митохондрия - двумембранная гранулярная или нитевидная органелла, элемент эукариотических клеток (автотрофов и гетеротрофов), энергетическая станция. Основная функция и энергообразование; происхождение, структура. Митохондриальная ДНК и наследственность.
Геном человека [Энциклопедия, написанная четырьмя буквами] Тарантул Вячеслав Залманович
25-я ХРОМОСОМА - ВАЖНОЕ ДОПОЛНЕНИЕ (митохондриальный геном)
Мал золотник, да дорог.
Русская поговорка
Когда сейчас громогласно заявляют о полном секвенировании генома человека, то, как правило, подразумевают ядерный геном. На этом фоне как-то забывается, что в клетках имеются молекулы ДНК, расположенные не только в хромосомах, но и в таких уже упоминавшихся специфических внутриклеточных структурах, как митохондрии. И это тоже геном человека, но он называется митохондриальным, а ДНК - митохондриальной (сокращенно митДНК). МитДНК теперь называют иногда хромосомой 25 или М-хромосомой. Эта ДНК была секвенирована еще в 1981 году уже упоминавшимся Ф. Сенгером, что тоже было в свое время сенсацией, которая, однако, имела резонанс несопоставимо меньший, чем секвенирование ядерного генома. Что же представляет собой эта 25-ая хромосома человека?
В клетке человека насчитывается от 100 до 1000 митохондрий, в каждой из которых содержится от 2 до 10 молекул кольцевой митДНК длиной 16569 п. н. Таким образом, размер митохондриального генома примерно в 200 000 раз меньше ядерного. Интересно, что размер митДНК у человека - один из наименьших среди высших организмов (эукариот). Например, у дрожжей митДНК состоит из 78520 п. н. Человеческая митДНК содержит 37 генов, кодирующих 13 белковых цепей, 22 тРНК и 2 рибосомных РНК (рРНК) (рис. 30). Белковые цепи входят в состав белков, которые участвуют в основном в важнейшем внутриклеточном процессе, называемом окислительным фосфорилированием, который обеспечивает клетку энергией. За счет окислительного фосфорилирования в митохондриях осуществляется производство более 90 % специальных молекул АТФ, являющихся основой энергетики клетки.
Рис. 30 . Структура митохондриального генома человека (митДНК). В митДНК содержится 22 гена, кодирующих тРНК, 2 рибосомных гена (16S и 12S рРНК) и 13 белок-кодирущих генов. Стрелками указано направление транскрипции генов. Сокращения: ND1-ND6, ND4L - гены субъединиц НАД-Н-дегидрогеназного комплекса; COI–COIII - гены субъединиц цитохром-с-оксидазы; ATP6, ATP8 - гены субъединиц АТФ-синтетазы; Cyt b - ген цитохрома b
Всего же в процессе окислительного фосфорилирования задействовано 87 генов, но все недостающие 74 кодируются не митохондриальным, а ядерным геномом. Интересно, что в ядерном геноме обнаруживаются участки, подобные митДНК. Предполагается, что в процессе эволюции и при различных патологиях имела место миграция части митДНК в ядерный геном.
Важно, что устройство митохондриального генома существенно отличается от ядерного. В первую очередь, для митДНК характерно очень компактное расположение генов, как и в геноме бактерий. В отличие от ядерного генома митохондриальные гены соседствуют друг с другом и между ними практически отсутствуют межгенные промежутки. В ряде случаев они даже перекрываются на один нуклеотид: последний нуклеотид одного гена оказывается первым в следующем за ним. То есть гены набиты в митохондриальной ДНК, как сельди в бочке. Кроме того, большинство митохондриальных генов не содержит такие характерные для ядерных генов структуры, как интроны. Но это еще не все отличия. Выяснилось, в частности, что митДНК не подвержена такой модификации, как метилирование, которая характерна для ядерной ДНК.
Однако особенное удивление исследователей вызвал генетических код, используемый в митДНК. Хотя генетический код универсален (за очень небольшим исключением) во всем живом мире, в митохондриях используется некий его необычный вариант. Большинство кодонов в митохондриальных генах сходны с теми, которые имеются в ядерной ДНК, но наряду с этим имеются и принципиальные отличия. В митДНК человека изменили свой смысл четыре кодона. Терминирующими стали кодоны АГА и АГГ. Кодон УГА, являющийся в ядерной ДНК терминирующим, в митДНК не только не вызывает остановки трансляции, а кодирует аминокислоту триптофан. Аминокислоту метионин кодируют не один кодон АУГ, а еще кодон АУА, который в ядерном геноме кодирует аминокислоту изолейцин.
МитДНК ответственна в клетке за синтез всего лишь нескольких митохондриальных белков. Но эти белки очень важны для клетки, поскольку участвуют в осуществлении одного из важнейших процессов - обеспечении клетки энергией. Таким образом, митДНК - весьма ценное приложение к Энциклопедии человека. Белки, кодируемые непосредственно генами митДНК, синтезируются тут же в митохондриях. Для этой цели используется собственная РНК-полимераза и собственный аппарат белкового синтеза. Причина ясна - генетический код митохондрий особый, нужна и особая система биосинтеза.
Далеко не все белки, которые нужны для автономного существования митохондрий, кодируются митохондриальным геномом и синтезируются здесь же. Для этого их геном слишком мал. Большая часть митохондриальных белков и отдельных субъединиц этих белков кодируется основным, т. е. ядерным геномом и синтезируется в цитоплазме клеток. Затем они транспортируются в митохондрии, где взаимодействуют со специфическими белками, кодируемыми митДНК. Таким образом, между ядерным и митохондриальным геномом существует тесная взаимосвязь, они дополняют друг друга.
Почему в эволюции клетки случилось так, что очень небольшая часть ДНК содержится не в хромосомах ядра, а отдельно внутри митохондрий? В чем необходимость или преимущество такого распределения генетического материала, пока неизвестно. Для объяснения этого удивительного факта было придумано много гипотез. Одну из первых еще в далеком 1890 году высказал Р. Альтман. Однако она и на сегодняшний день сохранила актуальность. Согласно этой точке зрения, митохондрии появились в клетках высших организмов не в ходе внутриклеточного развития и дифференцировки, а в результате естественного симбиоза высших организмов с низшими аэробными организмами. Такое объяснение предполагает, что митохондриальный генетический код более древен, чем код, используемый в ядерной ДНК у современных организмов.
Но наряду с этим высказывалась и иная точка зрения, которая пока в равной мере имеет право на существование. Согласно последней, после перехода большинства генов из митДНК в ядерную ДНК в аппарате, обеспечивающем синтез белка в митохондриях, произошли какие-то мутации. Для того, чтобы процесс трансляции не нарушался, потребовались специальные мутации в генах митДНК, которые бы «компенсировали» нарушения и позволили бы измененному аппарату белкового синтеза осуществлять свою работу. Если исходить из этого предположения, тогда митохондриальный код следует рассматривать не как более древний, а, наоборот, скорее как более молодой.
В любом случае язык митДНК - это в определенном смысл «жаргон». Зачем он нужен митохондриям? Здесь можно провести параллель с жаргонами определенных социальных или профессиональных групп. Жаргон используется ими для сокрытия своих намерений и действий от посторонних, избежания чужого вмешательства в их дела. Возможно, и митДНК, благодаря использованию видоизмененного кода - жаргона, - изолируется от белок-синтезирующиего аппарата клетки, специализируясь на выполнении одной, но очень важной для клетки функции - производстве энергии.
Замечено, что митохондриальный геном более раним, чем ядерный геном. В результате в нем часто происходят различного рода мутации (точковые мутации, небольшие потери ДНК - делеции и, наоборот, вставки - инсерции). Сейчас уже установлены многочисленные болезни человека, связанные с изменениями в митДНК. Патологические мутации обнаружены почти во всех митохондриальных генах. При этом отмечают огромное разнообразие клинических признаков, обусловленных одним и тем же молекулярным повреждением. Обнаружена взаимосвязь некоторых мутаций и изменений в экспрессии генов митДНК с возникновением рака. В частности, неоднократно отмечалось при раке груди и лимфомах усиление транскрипции гена, кодирующего одну из цепей белкого комплекса, участвующего в снабжении клеток энергией (субъединицы II цитохром-с - оксидазы). Некоторые, к счастью, редкие, тяжелые наследственные болезни человека также обусловлены мутациями в отдельных генах митДНК. В России сейчас существует специальная программа диагностики и профилактики митохондриальных болезней.
Еще один удивительный факт, связанный с митДНК, касается ее наследования. Оказалось, что митДНК передается из поколения в поколение принципиально иначе, чем хромосомная ДНК. Организм человека развивается из оплодотворенной яйцеклетки, которая содержит хромосомы обоих родителей. При оплодотворении в яйцо проникает сперматозоид с набором отцовских хромосом, но практически без отцовских митохондрий и, следовательно, без какой-либо отцовской митДНК. Только яйцеклетка предоставляет зародышу свою митДНК. Это ведет к важным последствиям: митДНК передается только по женской линии. Мы все получаем митДНК только от своей матери, а она еще раньше от своей, и так далее в ряду только женских поколений. Сыновья в отличие от дочерей не передают свою митДНК - цепочка оборвется. Таким путем ДНК образуют клоны - наследственные линии, которые могут только разветвляться (если у женщины родилось несколько дочерей), но в отличие от хромосомной ДНК не могут объединяться в одном организме и создавать новые генетические комбинации. По этой причине было интересно провести сравнение митДНК у представителей различных человеческих этнических популяций, то есть рас и народностей. Такого рода сравнения были начаты еще в конце 80-х годов прошлого века и продолжаются до сих пор. Подробнее мы еще поговорим об этом далее.
Таким образом, такие базовые процессы в клетке, как транскрипция, трансляция, репликация и репарация митДНК, в значительной мере зависят от ядерного генома, но пока не до конца ясно, как эти два генома интегрированы друг с другом. Изучение механизмов межгеномного взаимодействия может стать полезным во многих отношениях, в частности для понимания интегральной картины различных патологий человека, включая и злокачественное перерождение клеток.
Из книги ЧЕЛОВЕК - ты, я и первозданный автора Линдблад ЯнГлава 4 Наше самое важное ручное животное и его значение для возникновения земледелия. Примечательное развитие хлебных злаков. Ответить на вопрос, когда человек впервые приручил крупный рогатый скот, так же трудно, как на вопрос о времени одомашнивания других животных.
Из книги Гомеопатическое лечение кошек и собак автора Гамильтон ДонДополнение Словарь специальных терминов Аллопатия (alios - другой. pathos - болезнь). Метод лечения заболеваний по принципу противоположности. Врачи-аллопаты лечат болезнь, а не пациента. Этот термин обычно используется в отношении западной медицинской школы, которая
Из книги Геном человека: Энциклопедия, написанная четырьмя буквами автораЧТО ТАКОЕ ГЕНОМ? Вопросы вечны, ответы обусловлены временем. Е. Чаргафф В диалоге с жизнью важен не ее вопрос, а наш ответ. М. И. Цветаева С самого начала определимся, что мы здесь будем подразумевать под словом геном. Сам этот термин впервые был предложен в 1920 году немецким
Из книги Геном человека [Энциклопедия, написанная четырьмя буквами] автора Тарантул Вячеслав Залманович25-я ХРОМОСОМА - ВАЖНОЕ ДОПОЛНЕНИЕ (митохондриальный геном) Мал золотник, да дорог. Русская поговорка Когда сейчас громогласно заявляют о полном секвенировании генома человека, то, как правило, подразумевают ядерный геном. На этом фоне как-то забывается, что в клетках
Из книги Нерешенные проблемы теории эволюции автора Красилов Валентин АбрамовичЧТО ТАКОЕ ГЕНОМ? Вопросы вечны, ответы обусловлены временем. Е. Чаргафф В диалоге с жизнью важен не ее вопрос, а наш ответ. М. И. Цветаева С самого начала определимся, что мы здесь будем подразумевать под словом геном. Сам этот термин впервые был предложен в 1920 году
Из книги Виды психики: на пути к пониманию сознания автора Деннет ДэниэлВСЕ ЛИ В НАС ЗАВИСИТ ОТ ГЕНОМА? (геном и окружающая среда) Законам природы люди повинуются, даже когда борются против них. И. Гёте Как великий художник, природа умеет и с небольшими средствами достигать великих эффектов. Г. Гейне Конечно же, надо понимать, что
Из книги Удивительная палеонтология [История земли и жизни на ней] автора Еськов Кирилл Юрьевич Из книги Мир животных. Том 3 [Рассказы о птицах] автора Акимушкин Игорь ИвановичГлава 4. Как интенциональность приобрела важное
Из книги Эволюция человека. Книга 1. Обезьяны, кости и гены автора Марков Александр ВладимировичДОПОЛНЕНИЕ К ГЛАВЕ 4 Термодинамические подходы к сущности жизни. Второе начало термодинамики, энтропия и диссипативные структуры Нам придется начать издалека. В 1847 году Г. Гельмгольц сформулировал закон сохранения энергии (ЗСЭ). Следует помнить, что ЗСЭ является всего
Из книги В мире незримого автора Блинкин Семен АлександровичДОПОЛНЕНИЕ К ГЛАВЕ 6 Взаимоотношения хищника и жертвы в экологическом и эволюционном масштабах времени Одной из приоритетных задач современной биологии, несомненно, является создание обобщенной теории эволюции экосистем. При этом у экологов может возникнуть сильный
Из книги Расшифрованная жизнь [Мой геном, моя жизнь] автора Вентер КрейгДОПОЛНЕНИЕ К ГЛАВЕ 14 Историческая биогеография. Викариантная модель и концепция «оттесненных реликтов». Фитоспрединг. Биогеография - наука о закономерностях географического распространения живых существ. Вполне очевидно, что всякое живое существо экологически
Из книги Поведение: эволюционный подход автора Курчанов Николай АнатольевичДополнение к голенастым: отряд фламинго Фламинго голенаст, даже очень, – птица необыкновенно длинноногая. Но по резонным причинам, которые мы здесь обсуждать не будем, его из отряда голенастых (также из пластинчатоклювых, куда фламинго тоже зачисляли) ныне исключили,
Из книги автораГеном неандертальца Еще совсем недавно пределом мечтаний для палеогенетиков было выделение из древних костей митохондриальной ДНК. Эта небольшая часть генома, передающаяся по материнской линии, присутствует в каждой клетке в сотнях копий, к тому же она имеет
Из книги автораДополнение к пастеровским прививкам Новое и важное дополнение к пастеровским прививкам разработали ученые уже в XX в. Несколько лет назад советские ученые создали антирабический гамма-глобулин. С получением этого препарата предупреждение бешенства стало еще более
Из книги автораГлава 14 Первый геном человека Перспектива того, что тебя опередят в научной гонке, обычно вызывает отчаяние и безумную надежду – а вдруг повезет, и твой конкурент завтра помрет. Иной раз хочется просто все бросить, но тогда годы тяжелого труда будут потрачены
Из книги автора1.5. Лабильный геном Традиционные представления о стабильности геномов, сложившиеся в рамках классической генетики, сильно пошатнулись после открытия мобильных (мигрирующих) генетических элементов (МГЭ). МГЭ – это структуры, которые могут перемещаться в пределах генома
Управление